
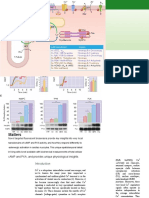
Wang 2019
Wang 2019
You might also like
- Toxins: The FST Potential Roles in Stress Response, Metabolism and PathogenesisDocument12 pagesToxins: The FST Potential Roles in Stress Response, Metabolism and Pathogenesismaría joséNo ratings yet
- GKW 1246Document12 pagesGKW 1246Thaís MesequeNo ratings yet
- 1 s2.0 S0021925819316795 MainDocument7 pages1 s2.0 S0021925819316795 MainLeidy Constanza Villalobos GonzalezNo ratings yet
- RNA Silencing: 90th International Titisee ConferenceDocument9 pagesRNA Silencing: 90th International Titisee ConferenceViệt KhoaNo ratings yet
- Calamar ArnmDocument14 pagesCalamar ArnmShin Chu Lan ShaNo ratings yet
- Kotov 2014 EnglDocument13 pagesKotov 2014 EnglLudmila OleninaNo ratings yet
- PMC4027209 Gku180Document10 pagesPMC4027209 Gku180Gotham BuddhaNo ratings yet
- 10 1002@bies 20085Document12 pages10 1002@bies 20085Minh Nguyen NgocNo ratings yet
- Dependence of ORC Silencing Function OnDocument13 pagesDependence of ORC Silencing Function Onkilli999No ratings yet
- E. Coli Has Been Thoroughly Studied in Gene RegulationDocument10 pagesE. Coli Has Been Thoroughly Studied in Gene Regulationdm_gty88No ratings yet
- Williams - 1999 - PKR A Sentinel Kinase For Cellular StressDocument9 pagesWilliams - 1999 - PKR A Sentinel Kinase For Cellular StressGuillem PratsNo ratings yet
- 77Document10 pages77karecuNo ratings yet
- Rab Proteins As Membrane Organizers: Marino Zerial and Heidi McbrideDocument13 pagesRab Proteins As Membrane Organizers: Marino Zerial and Heidi McbrideCourtney JohnsonNo ratings yet
- DNA Topology: Topoisomerases Keep It Simple: Andrew D. Bates and Anthony MaxwellDocument4 pagesDNA Topology: Topoisomerases Keep It Simple: Andrew D. Bates and Anthony MaxwellakibNo ratings yet
- 2017 Yasuda A Look at Plant Immunity Through The Window of The Multitasking Coreceptor BAK1Document9 pages2017 Yasuda A Look at Plant Immunity Through The Window of The Multitasking Coreceptor BAK1Raul DavilaNo ratings yet
- Protein Expression and Purification: Simone L. Pival, Ruth Birner-Gruenberger, Corinna Krump, Bernd NidetzkyDocument8 pagesProtein Expression and Purification: Simone L. Pival, Ruth Birner-Gruenberger, Corinna Krump, Bernd Nidetzkyade putriantiNo ratings yet
- Workshop 1Document13 pagesWorkshop 1DaisyNo ratings yet
- 01 - Doniselli Et Al. 2015Document14 pages01 - Doniselli Et Al. 2015Edgar Huerta CardenasNo ratings yet
- Nitrogen Fixing Cereals - How Close Are We? Nitrogen Fixing Cereals - How Close Are We?Document37 pagesNitrogen Fixing Cereals - How Close Are We? Nitrogen Fixing Cereals - How Close Are We?Vidushi Harsh GuptaNo ratings yet
- Xie 2015. Diacylglycerol Kinases in Membrane TraffickingDocument10 pagesXie 2015. Diacylglycerol Kinases in Membrane TraffickingmcrickboomNo ratings yet
- Pahala 1Document5 pagesPahala 1Daeng MangkaNo ratings yet
- Section 7C - Vesicle Tethering and Fusion F2023 - PDocument13 pagesSection 7C - Vesicle Tethering and Fusion F2023 - PWilliamNo ratings yet
- J. Biol. Chem.-1979-Bowman-2928-34Document8 pagesJ. Biol. Chem.-1979-Bowman-2928-34pankaj harinkhedeNo ratings yet
- Reelin Activates SRC Family Tyrosine Kinases in NeuronsDocument9 pagesReelin Activates SRC Family Tyrosine Kinases in NeuronsRam KNo ratings yet
- Biotech Reviewer 2nd Qtr. GRADE 8Document4 pagesBiotech Reviewer 2nd Qtr. GRADE 8Andrew P. Rico0% (1)
- Streptococcus Faecalis: Membrane Adenosine Triphosphatase FromDocument8 pagesStreptococcus Faecalis: Membrane Adenosine Triphosphatase FromSinta 2712No ratings yet
- Expression of A Drosophila Circadian ClockDocument12 pagesExpression of A Drosophila Circadian Clocklana valeryNo ratings yet
- Differential Gene Expression of ARGININE DECARBOXYLASE ADC1 and ADC2 in ArabidopsisDocument13 pagesDifferential Gene Expression of ARGININE DECARBOXYLASE ADC1 and ADC2 in ArabidopsismNo ratings yet
- RNA EditingDocument36 pagesRNA EditingzeynoleeeNo ratings yet
- Genome Editing: PLNT2530 (2020) Unit 9Document25 pagesGenome Editing: PLNT2530 (2020) Unit 9Swati JainNo ratings yet
- Import Routes and Nuclear Functions of Argonaute and Other Small RNA - Silencing ProteinsDocument12 pagesImport Routes and Nuclear Functions of Argonaute and Other Small RNA - Silencing ProteinsMariaAndreaLaraSalasNo ratings yet
- Neurosci Lett 2009Document5 pagesNeurosci Lett 2009wangyu81No ratings yet
- BJ - 1 s2.0 S0167488923000745 MainDocument3 pagesBJ - 1 s2.0 S0167488923000745 Mainserama15692uacNo ratings yet
- J. Biol. Chem.-2003-Ayyagari-1618-25Document8 pagesJ. Biol. Chem.-2003-Ayyagari-1618-25A. M. Mahedi HasanNo ratings yet
- SREBP Review InsectDocument10 pagesSREBP Review Insectaaki2627No ratings yet
- 2008 - Whole Genome PlatypusDocument11 pages2008 - Whole Genome Platypusj_wingerdNo ratings yet
- 6 Regulation of Gene ExpressionDocument49 pages6 Regulation of Gene ExpressionNaivedyam Vineeta MishraNo ratings yet
- Csir Net NotesDocument63 pagesCsir Net Notesssjusvinder75% (4)
- The Phosphate Makes A Difference Cellular Functions of NADPDocument10 pagesThe Phosphate Makes A Difference Cellular Functions of NADPYasif AbbasNo ratings yet
- The Many Pathways of RNA Degradation: Open ArchiveDocument24 pagesThe Many Pathways of RNA Degradation: Open ArchiveSakshi SharmaNo ratings yet
- Structure of LRRK1 and Mechanisms of Autoinhibition and ActivationDocument31 pagesStructure of LRRK1 and Mechanisms of Autoinhibition and Activation9868838836ankNo ratings yet
- The Molecular Basis of ApoptosisDocument42 pagesThe Molecular Basis of ApoptosisvictoryNo ratings yet
- Sirna and Mirna Processing: New Functions For Cajal Bodies: Olga Pontes and Craig S PikaardDocument7 pagesSirna and Mirna Processing: New Functions For Cajal Bodies: Olga Pontes and Craig S PikaardChristian Moises CasasNo ratings yet
- The Inflammasome in Pathogen Recognition and InflammationDocument6 pagesThe Inflammasome in Pathogen Recognition and InflammationSakshi IssarNo ratings yet
- NIH Public Access: The Polycomb Complex PRC2 and Its Mark in LifeDocument18 pagesNIH Public Access: The Polycomb Complex PRC2 and Its Mark in LifeAbi Wombat MendiolitaNo ratings yet
- Agarose GelsDocument8 pagesAgarose Gelsrapuenzal farooqiNo ratings yet
- Kvainickas 2017 - Retromer - and WASH-dependent Sorting of Nutrient Transporters Requires A Multivalent Interaction Network With ANKRD50Document19 pagesKvainickas 2017 - Retromer - and WASH-dependent Sorting of Nutrient Transporters Requires A Multivalent Interaction Network With ANKRD50Václav BočanNo ratings yet
- Parp 1 e 2Document6 pagesParp 1 e 2AnnalisaDorioNo ratings yet
- AMG2.5 Klein Replication 2018Document50 pagesAMG2.5 Klein Replication 2018Теодора ИвановаNo ratings yet
- Cre-Loxp Recombination For Genetic Modification: Presented byDocument18 pagesCre-Loxp Recombination For Genetic Modification: Presented byKIRTI SHARMANo ratings yet
- Review: Whole-Cell cAMP and PKA Activity Are Epiphenomena, Nanodomain SignalingDocument16 pagesReview: Whole-Cell cAMP and PKA Activity Are Epiphenomena, Nanodomain SignalingRenee DayNo ratings yet
- JMolBiol 2005 Fosmidomycin DXR StructureDocument13 pagesJMolBiol 2005 Fosmidomycin DXR StructureVincent GeruszNo ratings yet
- Topo IsomeraseDocument12 pagesTopo IsomeraseAcih AsihNo ratings yet
- Structures of HIV-1 RT-DNA Complexes Before and After Incorporation of The anti-AIDS Drug TenofovirDocument6 pagesStructures of HIV-1 RT-DNA Complexes Before and After Incorporation of The anti-AIDS Drug TenofovirRaras PramuditaNo ratings yet
- Nucleotides and Nucleic AcidsDocument68 pagesNucleotides and Nucleic AcidsVaibhav KumarNo ratings yet
- 5.2 Resolution and Detection of NADocument3 pages5.2 Resolution and Detection of NAALLISON PAMITTANNo ratings yet
- Proteolytic Dissection of Zab, The Z-DNA-binding Domain of Human ADAR1Document8 pagesProteolytic Dissection of Zab, The Z-DNA-binding Domain of Human ADAR1Saradindu GhoshNo ratings yet
- FullDocument7 pagesFullMouna RohanaNo ratings yet
- 5 Molecular Mechanisms of Gene RegulationDocument44 pages5 Molecular Mechanisms of Gene Regulationpramudi madushaniNo ratings yet
- XI Botany ChapterWise Prvs Questions HssliveDocument25 pagesXI Botany ChapterWise Prvs Questions Hsslivejithu63% (8)
- Pages From Watson Molecular Biology of The Gene 7th Edition c2014 Txtbk-2Document1 pagePages From Watson Molecular Biology of The Gene 7th Edition c2014 Txtbk-2Kunal SethNo ratings yet
- An Introduction To Genes and GenomesDocument13 pagesAn Introduction To Genes and GenomesLatif Ur RehmanNo ratings yet
- Dna Gel Electrophoresis: Gateway To Various Biotechnology InvestigationsDocument16 pagesDna Gel Electrophoresis: Gateway To Various Biotechnology InvestigationsNasrullah RoslanNo ratings yet
- 2.2-2.5 - MCQ AnswersDocument39 pages2.2-2.5 - MCQ AnswersMohammad Ali Al-IadyNo ratings yet
- Successful Application of CADDDocument16 pagesSuccessful Application of CADDnagendraNo ratings yet
- Test Bank For Biochemistry 7th Edition Jeremy M BergDocument6 pagesTest Bank For Biochemistry 7th Edition Jeremy M BergkhuyenhenryzwoowNo ratings yet
- Genbio2 Lesson 1 Plant and Animal Nutrition Part 1Document32 pagesGenbio2 Lesson 1 Plant and Animal Nutrition Part 1crystal janr macalisangNo ratings yet
- PCR MultiplexDocument2 pagesPCR MultiplexFatima VessaliusNo ratings yet
- Lab Notes For Module 567 BiochemDocument12 pagesLab Notes For Module 567 BiochemRexxie PabualanNo ratings yet
- A Level Chemistry Application Booklet 9701Document153 pagesA Level Chemistry Application Booklet 9701shahezaman_ali88% (24)
- Biotechnology and Its ApplicationDocument83 pagesBiotechnology and Its ApplicationKanmani SathiachandranNo ratings yet
- Male Sterility in Vegetables: June 2013Document10 pagesMale Sterility in Vegetables: June 2013Janardhan RaoNo ratings yet
- This Study Resource Was: Bromelain As An EnzymeDocument7 pagesThis Study Resource Was: Bromelain As An Enzymeapril hortilanoNo ratings yet
- Chapter 10 - Plant Metabolism Stern's Plant Biology Flashcards - QuizletDocument21 pagesChapter 10 - Plant Metabolism Stern's Plant Biology Flashcards - QuizletChristianAvelinoNo ratings yet
- Immunology in Haematology (Part 2)Document55 pagesImmunology in Haematology (Part 2)kiedd_04100% (4)
- Letran de Davao, IncDocument3 pagesLetran de Davao, IncPatricia DianeNo ratings yet
- Biology Coursework Enzymes ConclusionDocument4 pagesBiology Coursework Enzymes Conclusionafaydebwo100% (2)
- Cells PDFDocument107 pagesCells PDFida hadiNo ratings yet
- Protected Protein and Amino Acids in Ruminant Nutrition: January 2005Document6 pagesProtected Protein and Amino Acids in Ruminant Nutrition: January 2005Carolina RamónNo ratings yet
- The 1st International Drug Discovery WorkshopDocument60 pagesThe 1st International Drug Discovery WorkshopAhmed ElrashedyNo ratings yet
- EuglenaDocument5 pagesEuglenaAngel García MNo ratings yet
- Genetics Bits gpbr-111Document10 pagesGenetics Bits gpbr-111Adan FatimaNo ratings yet
- BME 553 Exam IDocument12 pagesBME 553 Exam Imgnieves11No ratings yet
- Lesson Plan PatrickDocument4 pagesLesson Plan PatrickPatrick Jerome SilvanoNo ratings yet
- Isicem 2018Document215 pagesIsicem 2018Mihai PopescuNo ratings yet
- Resistance Is Futile ... or Is It?: The Immune System and HIV InfectionDocument28 pagesResistance Is Futile ... or Is It?: The Immune System and HIV InfectionRachel Mwale MwazigheNo ratings yet
- Yohannes BirukDocument6 pagesYohannes BirukYohannes BirukNo ratings yet
- L3 Spec Adrenal2 2022 FinalDocument49 pagesL3 Spec Adrenal2 2022 FinalNona NanoNo ratings yet
- Transgenic GuidelineDocument12 pagesTransgenic GuidelineYogesh BanginwarNo ratings yet






















































































Download as pdf or txt
You might also like
- Toxins: The FST Potential Roles in Stress Response, Metabolism and PathogenesisDocument12 pagesToxins: The FST Potential Roles in Stress Response, Metabolism and Pathogenesismaría joséNo ratings yet
- GKW 1246Document12 pagesGKW 1246Thaís MesequeNo ratings yet
- 1 s2.0 S0021925819316795 MainDocument7 pages1 s2.0 S0021925819316795 MainLeidy Constanza Villalobos GonzalezNo ratings yet
- RNA Silencing: 90th International Titisee ConferenceDocument9 pagesRNA Silencing: 90th International Titisee ConferenceViệt KhoaNo ratings yet
- Calamar ArnmDocument14 pagesCalamar ArnmShin Chu Lan ShaNo ratings yet
- Kotov 2014 EnglDocument13 pagesKotov 2014 EnglLudmila OleninaNo ratings yet
- PMC4027209 Gku180Document10 pagesPMC4027209 Gku180Gotham BuddhaNo ratings yet
- 10 1002@bies 20085Document12 pages10 1002@bies 20085Minh Nguyen NgocNo ratings yet
- Dependence of ORC Silencing Function OnDocument13 pagesDependence of ORC Silencing Function Onkilli999No ratings yet
- E. Coli Has Been Thoroughly Studied in Gene RegulationDocument10 pagesE. Coli Has Been Thoroughly Studied in Gene Regulationdm_gty88No ratings yet
- Williams - 1999 - PKR A Sentinel Kinase For Cellular StressDocument9 pagesWilliams - 1999 - PKR A Sentinel Kinase For Cellular StressGuillem PratsNo ratings yet
- 77Document10 pages77karecuNo ratings yet
- Rab Proteins As Membrane Organizers: Marino Zerial and Heidi McbrideDocument13 pagesRab Proteins As Membrane Organizers: Marino Zerial and Heidi McbrideCourtney JohnsonNo ratings yet
- DNA Topology: Topoisomerases Keep It Simple: Andrew D. Bates and Anthony MaxwellDocument4 pagesDNA Topology: Topoisomerases Keep It Simple: Andrew D. Bates and Anthony MaxwellakibNo ratings yet
- 2017 Yasuda A Look at Plant Immunity Through The Window of The Multitasking Coreceptor BAK1Document9 pages2017 Yasuda A Look at Plant Immunity Through The Window of The Multitasking Coreceptor BAK1Raul DavilaNo ratings yet
- Protein Expression and Purification: Simone L. Pival, Ruth Birner-Gruenberger, Corinna Krump, Bernd NidetzkyDocument8 pagesProtein Expression and Purification: Simone L. Pival, Ruth Birner-Gruenberger, Corinna Krump, Bernd Nidetzkyade putriantiNo ratings yet
- Workshop 1Document13 pagesWorkshop 1DaisyNo ratings yet
- 01 - Doniselli Et Al. 2015Document14 pages01 - Doniselli Et Al. 2015Edgar Huerta CardenasNo ratings yet
- Nitrogen Fixing Cereals - How Close Are We? Nitrogen Fixing Cereals - How Close Are We?Document37 pagesNitrogen Fixing Cereals - How Close Are We? Nitrogen Fixing Cereals - How Close Are We?Vidushi Harsh GuptaNo ratings yet
- Xie 2015. Diacylglycerol Kinases in Membrane TraffickingDocument10 pagesXie 2015. Diacylglycerol Kinases in Membrane TraffickingmcrickboomNo ratings yet
- Pahala 1Document5 pagesPahala 1Daeng MangkaNo ratings yet
- Section 7C - Vesicle Tethering and Fusion F2023 - PDocument13 pagesSection 7C - Vesicle Tethering and Fusion F2023 - PWilliamNo ratings yet
- J. Biol. Chem.-1979-Bowman-2928-34Document8 pagesJ. Biol. Chem.-1979-Bowman-2928-34pankaj harinkhedeNo ratings yet
- Reelin Activates SRC Family Tyrosine Kinases in NeuronsDocument9 pagesReelin Activates SRC Family Tyrosine Kinases in NeuronsRam KNo ratings yet
- Biotech Reviewer 2nd Qtr. GRADE 8Document4 pagesBiotech Reviewer 2nd Qtr. GRADE 8Andrew P. Rico0% (1)
- Streptococcus Faecalis: Membrane Adenosine Triphosphatase FromDocument8 pagesStreptococcus Faecalis: Membrane Adenosine Triphosphatase FromSinta 2712No ratings yet
- Expression of A Drosophila Circadian ClockDocument12 pagesExpression of A Drosophila Circadian Clocklana valeryNo ratings yet
- Differential Gene Expression of ARGININE DECARBOXYLASE ADC1 and ADC2 in ArabidopsisDocument13 pagesDifferential Gene Expression of ARGININE DECARBOXYLASE ADC1 and ADC2 in ArabidopsismNo ratings yet
- RNA EditingDocument36 pagesRNA EditingzeynoleeeNo ratings yet
- Genome Editing: PLNT2530 (2020) Unit 9Document25 pagesGenome Editing: PLNT2530 (2020) Unit 9Swati JainNo ratings yet
- Import Routes and Nuclear Functions of Argonaute and Other Small RNA - Silencing ProteinsDocument12 pagesImport Routes and Nuclear Functions of Argonaute and Other Small RNA - Silencing ProteinsMariaAndreaLaraSalasNo ratings yet
- Neurosci Lett 2009Document5 pagesNeurosci Lett 2009wangyu81No ratings yet
- BJ - 1 s2.0 S0167488923000745 MainDocument3 pagesBJ - 1 s2.0 S0167488923000745 Mainserama15692uacNo ratings yet
- J. Biol. Chem.-2003-Ayyagari-1618-25Document8 pagesJ. Biol. Chem.-2003-Ayyagari-1618-25A. M. Mahedi HasanNo ratings yet
- SREBP Review InsectDocument10 pagesSREBP Review Insectaaki2627No ratings yet
- 2008 - Whole Genome PlatypusDocument11 pages2008 - Whole Genome Platypusj_wingerdNo ratings yet
- 6 Regulation of Gene ExpressionDocument49 pages6 Regulation of Gene ExpressionNaivedyam Vineeta MishraNo ratings yet
- Csir Net NotesDocument63 pagesCsir Net Notesssjusvinder75% (4)
- The Phosphate Makes A Difference Cellular Functions of NADPDocument10 pagesThe Phosphate Makes A Difference Cellular Functions of NADPYasif AbbasNo ratings yet
- The Many Pathways of RNA Degradation: Open ArchiveDocument24 pagesThe Many Pathways of RNA Degradation: Open ArchiveSakshi SharmaNo ratings yet
- Structure of LRRK1 and Mechanisms of Autoinhibition and ActivationDocument31 pagesStructure of LRRK1 and Mechanisms of Autoinhibition and Activation9868838836ankNo ratings yet
- The Molecular Basis of ApoptosisDocument42 pagesThe Molecular Basis of ApoptosisvictoryNo ratings yet
- Sirna and Mirna Processing: New Functions For Cajal Bodies: Olga Pontes and Craig S PikaardDocument7 pagesSirna and Mirna Processing: New Functions For Cajal Bodies: Olga Pontes and Craig S PikaardChristian Moises CasasNo ratings yet
- The Inflammasome in Pathogen Recognition and InflammationDocument6 pagesThe Inflammasome in Pathogen Recognition and InflammationSakshi IssarNo ratings yet
- NIH Public Access: The Polycomb Complex PRC2 and Its Mark in LifeDocument18 pagesNIH Public Access: The Polycomb Complex PRC2 and Its Mark in LifeAbi Wombat MendiolitaNo ratings yet
- Agarose GelsDocument8 pagesAgarose Gelsrapuenzal farooqiNo ratings yet
- Kvainickas 2017 - Retromer - and WASH-dependent Sorting of Nutrient Transporters Requires A Multivalent Interaction Network With ANKRD50Document19 pagesKvainickas 2017 - Retromer - and WASH-dependent Sorting of Nutrient Transporters Requires A Multivalent Interaction Network With ANKRD50Václav BočanNo ratings yet
- Parp 1 e 2Document6 pagesParp 1 e 2AnnalisaDorioNo ratings yet
- AMG2.5 Klein Replication 2018Document50 pagesAMG2.5 Klein Replication 2018Теодора ИвановаNo ratings yet
- Cre-Loxp Recombination For Genetic Modification: Presented byDocument18 pagesCre-Loxp Recombination For Genetic Modification: Presented byKIRTI SHARMANo ratings yet
- Review: Whole-Cell cAMP and PKA Activity Are Epiphenomena, Nanodomain SignalingDocument16 pagesReview: Whole-Cell cAMP and PKA Activity Are Epiphenomena, Nanodomain SignalingRenee DayNo ratings yet
- JMolBiol 2005 Fosmidomycin DXR StructureDocument13 pagesJMolBiol 2005 Fosmidomycin DXR StructureVincent GeruszNo ratings yet
- Topo IsomeraseDocument12 pagesTopo IsomeraseAcih AsihNo ratings yet
- Structures of HIV-1 RT-DNA Complexes Before and After Incorporation of The anti-AIDS Drug TenofovirDocument6 pagesStructures of HIV-1 RT-DNA Complexes Before and After Incorporation of The anti-AIDS Drug TenofovirRaras PramuditaNo ratings yet
- Nucleotides and Nucleic AcidsDocument68 pagesNucleotides and Nucleic AcidsVaibhav KumarNo ratings yet
- 5.2 Resolution and Detection of NADocument3 pages5.2 Resolution and Detection of NAALLISON PAMITTANNo ratings yet
- Proteolytic Dissection of Zab, The Z-DNA-binding Domain of Human ADAR1Document8 pagesProteolytic Dissection of Zab, The Z-DNA-binding Domain of Human ADAR1Saradindu GhoshNo ratings yet
- FullDocument7 pagesFullMouna RohanaNo ratings yet
- 5 Molecular Mechanisms of Gene RegulationDocument44 pages5 Molecular Mechanisms of Gene Regulationpramudi madushaniNo ratings yet
- XI Botany ChapterWise Prvs Questions HssliveDocument25 pagesXI Botany ChapterWise Prvs Questions Hsslivejithu63% (8)
- Pages From Watson Molecular Biology of The Gene 7th Edition c2014 Txtbk-2Document1 pagePages From Watson Molecular Biology of The Gene 7th Edition c2014 Txtbk-2Kunal SethNo ratings yet
- An Introduction To Genes and GenomesDocument13 pagesAn Introduction To Genes and GenomesLatif Ur RehmanNo ratings yet
- Dna Gel Electrophoresis: Gateway To Various Biotechnology InvestigationsDocument16 pagesDna Gel Electrophoresis: Gateway To Various Biotechnology InvestigationsNasrullah RoslanNo ratings yet
- 2.2-2.5 - MCQ AnswersDocument39 pages2.2-2.5 - MCQ AnswersMohammad Ali Al-IadyNo ratings yet
- Successful Application of CADDDocument16 pagesSuccessful Application of CADDnagendraNo ratings yet
- Test Bank For Biochemistry 7th Edition Jeremy M BergDocument6 pagesTest Bank For Biochemistry 7th Edition Jeremy M BergkhuyenhenryzwoowNo ratings yet
- Genbio2 Lesson 1 Plant and Animal Nutrition Part 1Document32 pagesGenbio2 Lesson 1 Plant and Animal Nutrition Part 1crystal janr macalisangNo ratings yet
- PCR MultiplexDocument2 pagesPCR MultiplexFatima VessaliusNo ratings yet
- Lab Notes For Module 567 BiochemDocument12 pagesLab Notes For Module 567 BiochemRexxie PabualanNo ratings yet
- A Level Chemistry Application Booklet 9701Document153 pagesA Level Chemistry Application Booklet 9701shahezaman_ali88% (24)
- Biotechnology and Its ApplicationDocument83 pagesBiotechnology and Its ApplicationKanmani SathiachandranNo ratings yet
- Male Sterility in Vegetables: June 2013Document10 pagesMale Sterility in Vegetables: June 2013Janardhan RaoNo ratings yet
- This Study Resource Was: Bromelain As An EnzymeDocument7 pagesThis Study Resource Was: Bromelain As An Enzymeapril hortilanoNo ratings yet
- Chapter 10 - Plant Metabolism Stern's Plant Biology Flashcards - QuizletDocument21 pagesChapter 10 - Plant Metabolism Stern's Plant Biology Flashcards - QuizletChristianAvelinoNo ratings yet
- Immunology in Haematology (Part 2)Document55 pagesImmunology in Haematology (Part 2)kiedd_04100% (4)
- Letran de Davao, IncDocument3 pagesLetran de Davao, IncPatricia DianeNo ratings yet
- Biology Coursework Enzymes ConclusionDocument4 pagesBiology Coursework Enzymes Conclusionafaydebwo100% (2)
- Cells PDFDocument107 pagesCells PDFida hadiNo ratings yet
- Protected Protein and Amino Acids in Ruminant Nutrition: January 2005Document6 pagesProtected Protein and Amino Acids in Ruminant Nutrition: January 2005Carolina RamónNo ratings yet
- The 1st International Drug Discovery WorkshopDocument60 pagesThe 1st International Drug Discovery WorkshopAhmed ElrashedyNo ratings yet
- EuglenaDocument5 pagesEuglenaAngel García MNo ratings yet
- Genetics Bits gpbr-111Document10 pagesGenetics Bits gpbr-111Adan FatimaNo ratings yet
- BME 553 Exam IDocument12 pagesBME 553 Exam Imgnieves11No ratings yet
- Lesson Plan PatrickDocument4 pagesLesson Plan PatrickPatrick Jerome SilvanoNo ratings yet
- Isicem 2018Document215 pagesIsicem 2018Mihai PopescuNo ratings yet
- Resistance Is Futile ... or Is It?: The Immune System and HIV InfectionDocument28 pagesResistance Is Futile ... or Is It?: The Immune System and HIV InfectionRachel Mwale MwazigheNo ratings yet
- Yohannes BirukDocument6 pagesYohannes BirukYohannes BirukNo ratings yet
- L3 Spec Adrenal2 2022 FinalDocument49 pagesL3 Spec Adrenal2 2022 FinalNona NanoNo ratings yet
- Transgenic GuidelineDocument12 pagesTransgenic GuidelineYogesh BanginwarNo ratings yet