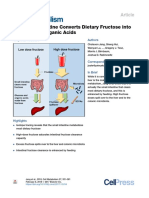
Download as pdf or txt
You might also like
- Harrison's Principles of Internal Medicine - 19e - Kasper, Fauci, Hauser, Longo, Jameson, Loscalzo (Dragged)Document16 pagesHarrison's Principles of Internal Medicine - 19e - Kasper, Fauci, Hauser, Longo, Jameson, Loscalzo (Dragged)Reg Lagarteja0% (1)
- Fructosa de La Dieta y Síndrome MetabólicoDocument16 pagesFructosa de La Dieta y Síndrome MetabólicoMariela RsNo ratings yet
- Fructose-Containing Caloric Sweeteners As A Cause of Obesity and Metabolic DisordersDocument9 pagesFructose-Containing Caloric Sweeteners As A Cause of Obesity and Metabolic DisordersJihan SMNo ratings yet
- 1 s2.0 S1550413117307295 MainDocument15 pages1 s2.0 S1550413117307295 MainGustavo AlmeidaNo ratings yet
- 1, Internship Report Tayyba 7th 2020.-1Document21 pages1, Internship Report Tayyba 7th 2020.-1S M Abrar HassanNo ratings yet
- Hormonios+e+Obesidade+-+Nutric A O+brasil+ (Science+Play)Document18 pagesHormonios+e+Obesidade+-+Nutric A O+brasil+ (Science+Play)Patrícia MedeirosNo ratings yet
- Bioavailability (Autosaved)Document7 pagesBioavailability (Autosaved)Sunu RodriguezNo ratings yet
- Skoog 2004Document5 pagesSkoog 2004özlemNo ratings yet
- Digestion and Absorption 10Document42 pagesDigestion and Absorption 10Pranjul MishraNo ratings yet
- Transport of Peptide and Protein Drugs Across BiologicalDocument11 pagesTransport of Peptide and Protein Drugs Across BiologicalJuan camilo Vargas perdonoNo ratings yet
- JA - 477 - Botezelli Et Al 2010Document8 pagesJA - 477 - Botezelli Et Al 2010jailtongpNo ratings yet
- A Review of Hereditary Fructose Intolerance: Tiberius Mogoș, Andra Evelin IacobiniDocument5 pagesA Review of Hereditary Fructose Intolerance: Tiberius Mogoș, Andra Evelin IacobiniKyla Barrera TabungarNo ratings yet
- Metformin LectureDocument12 pagesMetformin Lectureصادق التميميNo ratings yet
- Defectos MetabólicosDocument14 pagesDefectos Metabólicosandres herreraNo ratings yet
- Correcion Dosis Atb Falla RenalDocument6 pagesCorrecion Dosis Atb Falla RenalAMENDBENo ratings yet
- ESPEN Guidelines On Nutrition in Acute Pancreatitis: Consensus StatementDocument11 pagesESPEN Guidelines On Nutrition in Acute Pancreatitis: Consensus StatementMemo HadyNo ratings yet
- Drug Nmetabolism - BookDocument20 pagesDrug Nmetabolism - Bookcheonsa 1004No ratings yet
- Dietary Fructooligosaccharides and Potential Benefits On Health, Sabater-Molina 2009Document14 pagesDietary Fructooligosaccharides and Potential Benefits On Health, Sabater-Molina 2009manuel felipe villalbaNo ratings yet
- Nutrients 09 00385 Glicación AvanzDocument16 pagesNutrients 09 00385 Glicación AvanzLuis franNo ratings yet
- Phosphate A Poison For HumansDocument11 pagesPhosphate A Poison For HumansAssault AmphibiansNo ratings yet
- Digestive Enzyme Supplementation in Gastrointestinal DiseasesDocument7 pagesDigestive Enzyme Supplementation in Gastrointestinal Diseaseslewis.liusterNo ratings yet
- Digestion & Absorption ProteinsDocument44 pagesDigestion & Absorption ProteinsPutri Indah NirmalasariNo ratings yet
- Metabolism in Liver Kidney and PlacentaDocument22 pagesMetabolism in Liver Kidney and PlacentaBikash SahNo ratings yet
- 1 s2.0 S221287782030212X MainDocument17 pages1 s2.0 S221287782030212X MainAldoNo ratings yet
- Role of Digestive Tract Hormone in Functional Dyspepsia: Mohammad Adi Firmansyah, Dadang Makmun, Murdani AbdullahDocument5 pagesRole of Digestive Tract Hormone in Functional Dyspepsia: Mohammad Adi Firmansyah, Dadang Makmun, Murdani AbdullahYuneaHarisaNo ratings yet
- Obesity Gut MicrobesDocument12 pagesObesity Gut MicrobesBelindaDcosta0% (1)
- Glikogen 6Document12 pagesGlikogen 6DZoel FAcHMyNo ratings yet
- Prebiotics, Immune Function, Infection and in Ammation: A Review of The EvidenceDocument27 pagesPrebiotics, Immune Function, Infection and in Ammation: A Review of The EvidenceWahyu Ika WardhaniNo ratings yet
- Canfora 2019Document13 pagesCanfora 2019salamiNo ratings yet
- Fimmu 2017 01159Document17 pagesFimmu 2017 01159THAIS LENINE DE ALBUQUERQUENo ratings yet
- Review ArticleDocument10 pagesReview ArticlefransiscuNo ratings yet
- Biochemistry of Digestive SystemDocument17 pagesBiochemistry of Digestive SystemnishibuchiNo ratings yet
- Long-Term Fructose Consumption Accelerates Glycation and Several Age-Related Variables in Male RatsDocument8 pagesLong-Term Fructose Consumption Accelerates Glycation and Several Age-Related Variables in Male Ratsdumboo21No ratings yet
- GIT Lecture 2 Unit 3 2023Document5 pagesGIT Lecture 2 Unit 3 2023mashel JmilNo ratings yet
- 1 s2.0 S1756464615002121 MainDocument12 pages1 s2.0 S1756464615002121 MainDuzzysNo ratings yet
- Bioavailability of Flavonoids The Role of Cell Membrane TransportersDocument26 pagesBioavailability of Flavonoids The Role of Cell Membrane TransportersBRUNO JOSE BRITO TEIXEIRANo ratings yet
- Guo2012 PDFDocument10 pagesGuo2012 PDFLizbethNo ratings yet
- Metabolites 11 00479 PDFDocument23 pagesMetabolites 11 00479 PDFpond_1993No ratings yet
- MicroorganismsDocument21 pagesMicroorganismsDr LoloNo ratings yet
- 363 PDF PDFDocument7 pages363 PDF PDFEko Setyo BudiNo ratings yet
- MBBS Lecture 1 Gastrointestinal HormonesDocument5 pagesMBBS Lecture 1 Gastrointestinal Hormonessamueltoluwabori360No ratings yet
- Jurnal Kolon Spo 1 PDFDocument13 pagesJurnal Kolon Spo 1 PDFShenina KnightleyNo ratings yet
- 5 PDFDocument13 pages5 PDFAbu Reza A GaniNo ratings yet
- Fructosa y Su Relación Con Los CarbohidratosDocument10 pagesFructosa y Su Relación Con Los CarbohidratosLAURA CRISTINA CAÑON TRUJILLONo ratings yet
- Digestion Absorption 12 2019Document14 pagesDigestion Absorption 12 2019Karyan ShinNo ratings yet
- Gut Kidney 1Document10 pagesGut Kidney 1AlfirahmatikaNo ratings yet
- Carbohydrate Digestion NASPGHANDocument8 pagesCarbohydrate Digestion NASPGHANPradusha RevuruNo ratings yet
- Prescribing in Liver Disease Issue 2 Volume 32 Australian PrescriberDocument6 pagesPrescribing in Liver Disease Issue 2 Volume 32 Australian PrescriberAnonymous 9dVZCnTXSNo ratings yet
- Absorption Bioavailability and Metabolism of FlavonoidsDocument11 pagesAbsorption Bioavailability and Metabolism of FlavonoidsRena NormasariNo ratings yet
- Glucose Transporters in The 21st CenturyDocument9 pagesGlucose Transporters in The 21st CenturyThomas Edward Carreira FerreiraNo ratings yet
- tjp0591 0401Document14 pagestjp0591 0401özlemNo ratings yet
- Colon Targeted Drug DeliveryDocument21 pagesColon Targeted Drug DeliveryDeepak SharmaNo ratings yet
- Drug Interaction Linked To Drug TransporterDocument25 pagesDrug Interaction Linked To Drug TransporterPrarthna Yadav100% (1)
- The Exocrine Pancreas: Pancreatic Enzymes The Missing Link in Health and Disease?Document23 pagesThe Exocrine Pancreas: Pancreatic Enzymes The Missing Link in Health and Disease?Sunita RaniNo ratings yet
- Char Rez 2015Document11 pagesChar Rez 2015özlemNo ratings yet
- Vega-Paulino Potential Application of Commercial Enzyme Preparations For IndustrialDocument8 pagesVega-Paulino Potential Application of Commercial Enzyme Preparations For IndustrialRobertoNo ratings yet
- (14796813 - Journal of Molecular Endocrinology) Defining High-Fat-Diet Rat Models - Metabolic and Molecular Effects of Different Fat TypesDocument17 pages(14796813 - Journal of Molecular Endocrinology) Defining High-Fat-Diet Rat Models - Metabolic and Molecular Effects of Different Fat TypesNur Athirah HamzahNo ratings yet
- Microbial PathogenesisDocument8 pagesMicrobial PathogenesisRachelleNo ratings yet
- Gastrointestinal Tract and Digestion in The Young Ruminant: Ontogenesis, Adaptations, Consequences and ManipulationsDocument10 pagesGastrointestinal Tract and Digestion in The Young Ruminant: Ontogenesis, Adaptations, Consequences and ManipulationsSusila Semadi PutraNo ratings yet
- On Digestive Proteolysis: Being the Cartwright Lectures for 1894From EverandOn Digestive Proteolysis: Being the Cartwright Lectures for 1894No ratings yet
- Cholesterol Liquicolor CHOD-PAPDocument1 pageCholesterol Liquicolor CHOD-PAPmaarboleda1100% (2)
- Blood ClottingDocument11 pagesBlood ClottingsimikuttiyaniNo ratings yet
- Nursing Research Paper FinalDocument15 pagesNursing Research Paper Finalapi-546266464No ratings yet
- Laboratory Normal Reference Ranges: Children's Hospital and Health Center - San DiegoDocument6 pagesLaboratory Normal Reference Ranges: Children's Hospital and Health Center - San Diegoوليد. وفي الروحNo ratings yet
- Case Study PresentationDocument35 pagesCase Study PresentationblairNo ratings yet
- Dyslipidemia: Annals of Internal MedicineDocument16 pagesDyslipidemia: Annals of Internal MedicineCalvin LitraNo ratings yet
- Star Health and Allied Insurance Company Limited Tariff Statement - Mini SOC (FINALISED)Document1 pageStar Health and Allied Insurance Company Limited Tariff Statement - Mini SOC (FINALISED)Sanket Sahare100% (1)
- Access 25 (OH) Vitamin D Assay Total IFUDocument16 pagesAccess 25 (OH) Vitamin D Assay Total IFUcarineNo ratings yet
- Guidelines For Management of Dyslipidemia and Prevention of Cardiovascular DiseaseDocument78 pagesGuidelines For Management of Dyslipidemia and Prevention of Cardiovascular DiseasesabaxlentNo ratings yet
- Endorsement LetterDocument1 pageEndorsement LetterBernard CasimiroNo ratings yet
- HDL Cholesterol: HDLC - PDocument2 pagesHDL Cholesterol: HDLC - PMariiCarmen Ferez FeraldhNo ratings yet
- Valores de Laboratorio Harriet LaneDocument14 pagesValores de Laboratorio Harriet LaneRonald MoralesNo ratings yet
- Lipid Chemistry and MetabolismDocument4 pagesLipid Chemistry and Metabolism20AR018 HARIHARA SUBRAMANIANNo ratings yet
- Diabetes Leader 2019 PDFDocument12 pagesDiabetes Leader 2019 PDFArjun Kumar AdhikariNo ratings yet
- Department of Chemical Pathology: Lipid ProfileDocument2 pagesDepartment of Chemical Pathology: Lipid ProfileShahab KhanNo ratings yet
- Rogando-Santos, Eleni Tan 2245023050Document3 pagesRogando-Santos, Eleni Tan 2245023050Darby SantosNo ratings yet
- The Journal For Nurse Practitioners: Jacqueline O'Toole, MSC, NP, Irene Gibson, Ma, RGN, Gerard T. Flaherty, MDDocument4 pagesThe Journal For Nurse Practitioners: Jacqueline O'Toole, MSC, NP, Irene Gibson, Ma, RGN, Gerard T. Flaherty, MDGina GuisamanoNo ratings yet
- 3 Statins (Atorvastatin & Rosuvastatin)Document15 pages3 Statins (Atorvastatin & Rosuvastatin)Thet Su LwinNo ratings yet
- Group1 Bipolar DisorderDocument145 pagesGroup1 Bipolar DisorderEula Angelica OcoNo ratings yet
- Lecithin Book by Carlson WadeDocument195 pagesLecithin Book by Carlson WadeYoBjjNo ratings yet
- Mia Schow Consumer Interest Article FinalDocument4 pagesMia Schow Consumer Interest Article Finalapi-741989823No ratings yet
- Steroid PathwaysDocument1 pageSteroid PathwaysCalvin ChiuNo ratings yet
- Limitations of Conventional Methods of Self-Monitoring of Blood GlucoseDocument5 pagesLimitations of Conventional Methods of Self-Monitoring of Blood GlucoseMohammed ElnaggarNo ratings yet
- HTH668 Chp.6 - Hospitality Strategic ManagementDocument5 pagesHTH668 Chp.6 - Hospitality Strategic ManagementDanial D'Haqim M.NNo ratings yet
- Blood ChemistryDocument3 pagesBlood ChemistryJennifer MalunaoNo ratings yet
- Probiotic 13 Fr-2021-845 Wulandari Aprl 2023Document11 pagesProbiotic 13 Fr-2021-845 Wulandari Aprl 2023wisnuadi UMBYNo ratings yet
- Fact Sheeton Canola OilDocument15 pagesFact Sheeton Canola OilMonika ThadeaNo ratings yet
- Jurnal Inter Imun Dan Lemak 1Document5 pagesJurnal Inter Imun Dan Lemak 1Regitaand PutNo ratings yet
- Shiela May Santillan Mejorada 142 Prk. Santol, Mandalagan, Bacolod City, Negros Occidental, Philippines Position Desired: Medical TechnologistDocument7 pagesShiela May Santillan Mejorada 142 Prk. Santol, Mandalagan, Bacolod City, Negros Occidental, Philippines Position Desired: Medical Technologistmejorada100% (1)