Download as pdf or txt
You might also like
- Ingber Lecture: From Cellular Mechanotransduction To Biologically Inspired EngineeringDocument14 pagesIngber Lecture: From Cellular Mechanotransduction To Biologically Inspired EngineeringTensegrity Wiki100% (2)
- Biology 20 Unit A TableDocument4 pagesBiology 20 Unit A Tableapi-236289588No ratings yet
- Lab 2Document11 pagesLab 2Han Hui Chong0% (1)
- Articulo. La Termodinámica Del Desarrollo en El Diseño de Tejidos BioartificialesDocument11 pagesArticulo. La Termodinámica Del Desarrollo en El Diseño de Tejidos BioartificialesFernanda DíazNo ratings yet
- Life - PaperDocument32 pagesLife - PaperPrashant VemuriNo ratings yet
- Self-Organized Cytoskeletal Alignment During Drosophila Mesoderm InvaginationDocument6 pagesSelf-Organized Cytoskeletal Alignment During Drosophila Mesoderm InvaginationVasillis MamosNo ratings yet
- A Computational Model of Organism Development and Carcinogenesis Resulting From Cells' Bioelectric Properties and CommunicationDocument14 pagesA Computational Model of Organism Development and Carcinogenesis Resulting From Cells' Bioelectric Properties and CommunicationAlan de OliveiraNo ratings yet
- 2020 - Biomimetic Peptide Self Assembly For Functional MaterialsDocument20 pages2020 - Biomimetic Peptide Self Assembly For Functional MaterialsdymscientificNo ratings yet
- Synthetic Biology: Steven A. Benner and A. Michael SismourDocument11 pagesSynthetic Biology: Steven A. Benner and A. Michael SismourGanesh NagarajanNo ratings yet
- Evolutionary Game Theory - Cells As PlayersDocument22 pagesEvolutionary Game Theory - Cells As PlayersMaria GianfredaNo ratings yet
- BioEssays - 2002 - Tyson - The Dynamics of Cell Cycle RegulationDocument15 pagesBioEssays - 2002 - Tyson - The Dynamics of Cell Cycle RegulationNindya AgustiaNo ratings yet
- An Approach To The de Novo Synthesis of LifeDocument11 pagesAn Approach To The de Novo Synthesis of LifeJohnNo ratings yet
- Morphogenetic Fields in Embryogenesis, Regeneration, and Cancer - Non-Local Control of Complex PatterningDocument41 pagesMorphogenetic Fields in Embryogenesis, Regeneration, and Cancer - Non-Local Control of Complex PatterningritaNo ratings yet
- Matriz Evaluativa Primer Periodo de BiologíaDocument2 pagesMatriz Evaluativa Primer Periodo de BiologíagabrielaNo ratings yet
- Artificial Cells From Basic Science To ApplicationDocument17 pagesArtificial Cells From Basic Science To ApplicationNo Speed LimitNo ratings yet
- Trends in Cell Biology - December 2012Document83 pagesTrends in Cell Biology - December 2012Scientific Publication BDNo ratings yet
- From Self Vitality System To Well Coordinated Patterns I Neuro Circulatory PerfusionDocument14 pagesFrom Self Vitality System To Well Coordinated Patterns I Neuro Circulatory PerfusionHerald Scholarly Open AccessNo ratings yet
- The Evolution of Molecular Biology Into Systems Biology: Nature Biotechnology November 2004Document5 pagesThe Evolution of Molecular Biology Into Systems Biology: Nature Biotechnology November 2004Gia HoàngNo ratings yet
- Definición Sistema BiólogicoDocument7 pagesDefinición Sistema BiólogicoDany NahomiNo ratings yet
- Milestones and Current Achievements in Development of Multif 2021 BioactiveDocument27 pagesMilestones and Current Achievements in Development of Multif 2021 BioactiveDrago MirNo ratings yet
- CH 2 BiologyDocument74 pagesCH 2 BiologyParkash Kumar RathoreNo ratings yet
- TRENDS in Biotechnology Vol.20 No.1 January 2002Document6 pagesTRENDS in Biotechnology Vol.20 No.1 January 2002gopikrrishna.jNo ratings yet
- Biology Grade 10 Unit PlanDocument4 pagesBiology Grade 10 Unit PlanNicketa AndersonNo ratings yet
- Dna and The Chemistry of Inheritance - Barry CommonerDocument25 pagesDna and The Chemistry of Inheritance - Barry CommonerLuisa MoonNo ratings yet
- Is The Cell Really A MachineDocument19 pagesIs The Cell Really A Machinezune153No ratings yet
- Jerka Et Al 2024 Unraveling Endothelial Cell Migration Insights Into Fundamental Forces Inflammation BiomaterialDocument16 pagesJerka Et Al 2024 Unraveling Endothelial Cell Migration Insights Into Fundamental Forces Inflammation Biomaterialdfkm5mjn7vNo ratings yet
- Following The Organism To Map Synthetic GenomicsDocument4 pagesFollowing The Organism To Map Synthetic GenomicsGRAÇAS PERFUMARIANo ratings yet
- The Long Road To A Synthetic Self-Replicating Central Dogma: Biochemistry March 2023Document13 pagesThe Long Road To A Synthetic Self-Replicating Central Dogma: Biochemistry March 2023Qwyn Kym De GuzmanNo ratings yet
- Perspectives: A Road Map For The Development of Community Systems (Cosy) BiologyDocument7 pagesPerspectives: A Road Map For The Development of Community Systems (Cosy) BiologyandreaNo ratings yet
- Unit 2 - Introduction To BiochemistryDocument8 pagesUnit 2 - Introduction To BiochemistryWinrich Louise M. MontanoNo ratings yet
- SSGBIO1 - CM Week 1Document16 pagesSSGBIO1 - CM Week 1sgdick31No ratings yet
- Collective Cell Migration: FurtherDocument25 pagesCollective Cell Migration: FurtherFrancesco TorelliNo ratings yet
- Human On ChipDocument14 pagesHuman On ChipYong SiewkuanNo ratings yet
- Kinetics of Substrate Utilization, Product Formation and BiomassDocument25 pagesKinetics of Substrate Utilization, Product Formation and BiomassVINCE VITRIOLONo ratings yet
- Evolution of Intracellular CompartmentalizationDocument13 pagesEvolution of Intracellular CompartmentalizationDiana MurgaNo ratings yet
- Anatomy and Physiology NotesDocument12 pagesAnatomy and Physiology Notesarnienina.niloNo ratings yet
- PhysiomeDocument4 pagesPhysiomesuman palNo ratings yet
- The Plasticity of Biocompatibility - 2023 - BiomaterialsDocument27 pagesThe Plasticity of Biocompatibility - 2023 - BiomaterialsCabbagePotatoNo ratings yet
- Biochem #1Document28 pagesBiochem #1Christian Jay AlvarezNo ratings yet
- BEST - BCL - 1 - 2 - Key Concept - Cells and Cell StructuresDocument5 pagesBEST - BCL - 1 - 2 - Key Concept - Cells and Cell StructuresmtauhidNo ratings yet
- Course Outline FUNCORE 102 General Zoology 1st Sem 2022-2023Document5 pagesCourse Outline FUNCORE 102 General Zoology 1st Sem 2022-2023Gracia NicolaiNo ratings yet
- Reciprocal LinkageDocument14 pagesReciprocal LinkageGirdhari Lal SainiNo ratings yet
- Be 1Document37 pagesBe 1rohit adhikaryNo ratings yet
- Advanced NanoBiomed Research - 2021 - Ratri - Recent Advances in Regenerative Tissue Fabrication Tools Materials andDocument20 pagesAdvanced NanoBiomed Research - 2021 - Ratri - Recent Advances in Regenerative Tissue Fabrication Tools Materials andKalaaji WirasataNo ratings yet
- Omplex Emergence and The Living Organization AN Epistemological Framework For BiologyDocument27 pagesOmplex Emergence and The Living Organization AN Epistemological Framework For Biologyngonzalezduran5920No ratings yet
- Animal Cell Parts and Functions: Organelle Summary of FunctionDocument4 pagesAnimal Cell Parts and Functions: Organelle Summary of FunctionDorothy Mayo MejesNo ratings yet
- Electroactive Biomaterials and Systems For Cell Fate Determination and Tissue Regeneration: Design and ApplicationsDocument33 pagesElectroactive Biomaterials and Systems For Cell Fate Determination and Tissue Regeneration: Design and ApplicationsmahaNo ratings yet
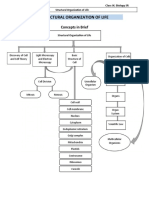
- Levels of Biological OrganizationDocument6 pagesLevels of Biological OrganizationRicardo RicoNo ratings yet
- EpidemiologyDocument21 pagesEpidemiologyAgustina TutiNo ratings yet
- The Evolution of Molecular Biology Into Systems BiologyDocument4 pagesThe Evolution of Molecular Biology Into Systems BiologyJuan Pablo Juárez CastilloNo ratings yet
- 1.cell Mechanics Principles Practices and ProspectsDocument18 pages1.cell Mechanics Principles Practices and ProspectsAlvaro Cano TortajadaNo ratings yet
- Advanced Materials - 2019 - Steier - Emerging Trends in Information Driven Engineering of Complex Biological SystemsDocument22 pagesAdvanced Materials - 2019 - Steier - Emerging Trends in Information Driven Engineering of Complex Biological SystemsramaNo ratings yet
- 2 Concept - About CellsDocument10 pages2 Concept - About CellskalloliNo ratings yet
- Fox Keller ClashOfTwoCulturesDocument2 pagesFox Keller ClashOfTwoCulturesFernando Adrian ZanczukNo ratings yet
- Cell Definition, Types, & Functions BritannicaDocument10 pagesCell Definition, Types, & Functions BritannicaKolade YousuffNo ratings yet
- Lesson 1 Characteristics of LifeDocument5 pagesLesson 1 Characteristics of LifeKrish D.No ratings yet
- 5 5 Chapter 75. Biofabricated Three Dimensional Tissue ModelsDocument25 pages5 5 Chapter 75. Biofabricated Three Dimensional Tissue ModelsblixesssNo ratings yet
- Abstract Biological Relativity and Circular Causality NobleDocument12 pagesAbstract Biological Relativity and Circular Causality NobleDaniel Felipe Marin VanegasNo ratings yet
- Thinking: BiologyDocument7 pagesThinking: BiologyJohn Paul VillabanNo ratings yet
- Self-Organization in Biological SystemsFrom EverandSelf-Organization in Biological SystemsRating: 4.5 out of 5 stars4.5/5 (7)
- Biology Unleashed: A Comprehensive Guide to Mastering the Science of LifeFrom EverandBiology Unleashed: A Comprehensive Guide to Mastering the Science of LifeNo ratings yet
- Preventing Chronic Disease:: Physical Activity and Healthy EatingDocument54 pagesPreventing Chronic Disease:: Physical Activity and Healthy EatingCHANGEZ KHAN SARDARNo ratings yet
- Scientists ListDocument7 pagesScientists ListsaniyasbaigNo ratings yet
- Photosynthesis Presentation NXPowerLiteDocument23 pagesPhotosynthesis Presentation NXPowerLite3DSツTRĪCKSHØTNo ratings yet
- Ranitidine (Zantac)Document3 pagesRanitidine (Zantac)sshah56No ratings yet
- Tittle: Human As A Complex OrganismDocument21 pagesTittle: Human As A Complex OrganismNick LeeNo ratings yet
- Sex Reversal in FishDocument5 pagesSex Reversal in FishKgerbNo ratings yet
- Cytotoxicity of GuttaFlow Bioseal, GuttaFlow2, MTA Fillapex, and AH Plus On Human Periodontal Ligament Stem CellsDocument7 pagesCytotoxicity of GuttaFlow Bioseal, GuttaFlow2, MTA Fillapex, and AH Plus On Human Periodontal Ligament Stem CellschristianNo ratings yet
- General Biology 1 Workbook Pages 43 52Document12 pagesGeneral Biology 1 Workbook Pages 43 52R VHINo ratings yet
- Bufadenolid 3Document239 pagesBufadenolid 3Diah Ayu WulandariNo ratings yet
- Bio 420 Final PresentationDocument11 pagesBio 420 Final Presentationapi-610305956No ratings yet
- Laboratory Activity 4Document4 pagesLaboratory Activity 4Sabrina MascardoNo ratings yet
- Lab Investigation AP 3 - Constructing Evol Lineages With DNA BLASTDocument7 pagesLab Investigation AP 3 - Constructing Evol Lineages With DNA BLASTKeri Gobin SamarooNo ratings yet
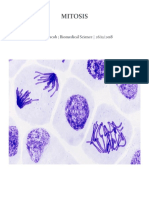
- MitosisDocument10 pagesMitosisAnonymous VS9jKQR0g0% (1)
- Inborn Errors of Metabolism IEM 1Document36 pagesInborn Errors of Metabolism IEM 1bhaskar chinnanandhi100% (1)
- David de Veredicis 0603298x - Mscmed Dissertation - Humg8006 - 14112016Document158 pagesDavid de Veredicis 0603298x - Mscmed Dissertation - Humg8006 - 14112016Abongile PhinyanaNo ratings yet
- The Balancing Act - Immunology of LeishmaniosisDocument7 pagesThe Balancing Act - Immunology of LeishmaniosisLaís Milagres BarbosaNo ratings yet
- HISTOPATH Introduction PDFDocument100 pagesHISTOPATH Introduction PDFpuja prajapatiNo ratings yet
- (L-2) - Cockroach - Jan 29, 2020Document45 pages(L-2) - Cockroach - Jan 29, 2020Harshit Sharma100% (1)
- Biosafety Biosecurity Biorisk Mngt.Document31 pagesBiosafety Biosecurity Biorisk Mngt.LawrenceNo ratings yet
- Bio HL Retake ExamDocument26 pagesBio HL Retake ExamHabiba Rafaat HamedNo ratings yet
- Cleavage, Blastula, Gastrula, NeurulaDocument41 pagesCleavage, Blastula, Gastrula, NeurulaHyenaNo ratings yet
- G. Muscle Physiology - 1Document45 pagesG. Muscle Physiology - 1Milimo MweembaNo ratings yet
- Assignment No. 1 Historical Antecedents in The WorldDocument5 pagesAssignment No. 1 Historical Antecedents in The Worldrommel benamirNo ratings yet
- The p63 Gene in EEC and Other Syndromes: Review ArticleDocument6 pagesThe p63 Gene in EEC and Other Syndromes: Review ArticleMishary Saud AlmalkeyNo ratings yet
- Certificate of Analysis: Test Items Specifications ResultsDocument1 pageCertificate of Analysis: Test Items Specifications ResultsCretulescu CameliaNo ratings yet
- 2017 SPM BIOLOGY Trial PaperDocument10 pages2017 SPM BIOLOGY Trial PaperV. SundrramNo ratings yet
- AFMC Question Paper BiologyDocument4 pagesAFMC Question Paper BiologySandeep MalakerNo ratings yet
- 50 Items Multiple ChoiceDocument8 pages50 Items Multiple ChoiceJueren MaciaNo ratings yet