Download as pdf or txt
You might also like
- Microbes and the Mind: The Impact of the Microbiome on Mental HealthFrom EverandMicrobes and the Mind: The Impact of the Microbiome on Mental HealthNo ratings yet
- Affidavit of Discrepancy Date of BirthDocument1 pageAffidavit of Discrepancy Date of Birthdempearl2315100% (1)
- APSR-17.7Annual Verification CertificateDocument2 pagesAPSR-17.7Annual Verification CertificatekkkothaNo ratings yet
- Tata Motors StrategyDocument17 pagesTata Motors StrategyRituraj Sen67% (3)
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument16 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsIqbal PrawiraNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument22 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsChris NeophytouNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument21 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistspuspitaNo ratings yet
- Escherichia Coli As A Model Organism and Its Application in Biotechnology PDFDocument23 pagesEscherichia Coli As A Model Organism and Its Application in Biotechnology PDFPerla AlvaradoNo ratings yet
- Crosstalk Between The Microbiota-Gut-Brain Axis and DepressionDocument8 pagesCrosstalk Between The Microbiota-Gut-Brain Axis and DepressionCamila VasquesNo ratings yet
- Metabolomics Basic Principles and StrategiesDocument16 pagesMetabolomics Basic Principles and StrategiesEmiliano TorresNo ratings yet
- Detection of Nutrient-Related SNP To Reveal Individual Malnutrition RiskDocument15 pagesDetection of Nutrient-Related SNP To Reveal Individual Malnutrition RiskBoo omNo ratings yet
- Chronobiology of Acid-Base Balance Under General A PDFDocument39 pagesChronobiology of Acid-Base Balance Under General A PDFM Fathur Arief KurniawanNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument39 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistssumanchyNo ratings yet
- Spielman Et Al., 2018-Unhealthy Gut, Unhealthy Brain - The Role of The Intestinal Microbiota in Neurodegenerative DiseasesDocument15 pagesSpielman Et Al., 2018-Unhealthy Gut, Unhealthy Brain - The Role of The Intestinal Microbiota in Neurodegenerative DiseasesEdwar MagallanNo ratings yet
- The Challenging Triad Microbiota ImmuneDocument15 pagesThe Challenging Triad Microbiota ImmuneJULIETANo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument20 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsBüşra YılmazNo ratings yet
- TFG Elisa AlósDocument38 pagesTFG Elisa AlósTuna TunNo ratings yet
- Investigating Gut-Brain Interaction: Connection Between Gut Microbes and Neurological HealthDocument8 pagesInvestigating Gut-Brain Interaction: Connection Between Gut Microbes and Neurological HealthInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Ijms 22 10028Document19 pagesIjms 22 10028daniela altamiranoNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument41 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsOMiD QahqaieNo ratings yet
- 52 - What S Bugging The Gut in Ocd A Review of The Gut Microbiome in ObsessiveDocument8 pages52 - What S Bugging The Gut in Ocd A Review of The Gut Microbiome in ObsessivePaula CantalapiedraNo ratings yet
- Relationship Between The Gut Microbiome and Brain Function RGDocument16 pagesRelationship Between The Gut Microbiome and Brain Function RGNaved MalikNo ratings yet
- TLTK Đ Án LectinDocument27 pagesTLTK Đ Án LectinPhuong TranNo ratings yet
- The Progress of Gut Microbiome Research Related To Brain DisordersDocument20 pagesThe Progress of Gut Microbiome Research Related To Brain DisordersVictoria HristovaNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument15 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsSaad MaqboolNo ratings yet
- Neuroscience and Biobehavioral Reviews: Bogdana Golofast, Karel Vales TDocument20 pagesNeuroscience and Biobehavioral Reviews: Bogdana Golofast, Karel Vales Ttimea_gheneaNo ratings yet
- Chapitre Sur Les PrebiotiquesDocument23 pagesChapitre Sur Les PrebiotiquesrachaNo ratings yet
- Microbial Endocrinology - The Microbiota - Gut-Brain Axis in Health and DiseaseDocument440 pagesMicrobial Endocrinology - The Microbiota - Gut-Brain Axis in Health and DiseaseCristiane ResendeNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument13 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsBüşra YılmazNo ratings yet
- Psychobiotics: A Novel Class of Psychotropic: EviewDocument7 pagesPsychobiotics: A Novel Class of Psychotropic: Eviewdavid cortezNo ratings yet
- 2013.11 Psicobióticos. Una Nueva Clase de PsicotrópicosDocument7 pages2013.11 Psicobióticos. Una Nueva Clase de PsicotrópicosEdward UriarteNo ratings yet
- Jurnal Pre-ProbiotikDocument10 pagesJurnal Pre-ProbiotikRistania96No ratings yet
- Artigos IDocument618 pagesArtigos IDanielMoraisMedNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument18 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsmusicontheroaddNo ratings yet
- Gut Microbiota in Human Metabolic Health and Disease: Yong Fan and Oluf PedersenDocument17 pagesGut Microbiota in Human Metabolic Health and Disease: Yong Fan and Oluf PedersenEsthéfane DarsNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument21 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsUnnati GargNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument18 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsBüşra YılmazNo ratings yet
- Eje Intestino-Cerebro-Microbioma.Document41 pagesEje Intestino-Cerebro-Microbioma.cristinaNo ratings yet
- Gut Flora in Health and DiseaseDocument8 pagesGut Flora in Health and DiseaseJohnny AtmanNo ratings yet
- Hepatocytes and Its Role in Metabolism12Document17 pagesHepatocytes and Its Role in Metabolism12cetinaNo ratings yet
- The Potential of Human Milk Oligosaccharides To Impact The Microbiota-Gut-Brain Axis Through Modulation of The Gut MicrobiotaDocument13 pagesThe Potential of Human Milk Oligosaccharides To Impact The Microbiota-Gut-Brain Axis Through Modulation of The Gut MicrobiotackrodriguesNo ratings yet
- Depressão e MicrobiomaDocument14 pagesDepressão e MicrobiomaDoc Nelson JuniorNo ratings yet
- Transferring The Blues Depression Associated Gut Microbiota Induces Neurobehavioural Changes in The Rat.Document11 pagesTransferring The Blues Depression Associated Gut Microbiota Induces Neurobehavioural Changes in The Rat.Dori BearNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument10 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsRazia UrmiNo ratings yet
- Porcine Islet Cell Xenotransplantation PDFDocument24 pagesPorcine Islet Cell Xenotransplantation PDFLemour YousefNo ratings yet
- Exercise Diet and Stress As Modulators of Gut Microbiota - 2020 - NeurobiologyDocument16 pagesExercise Diet and Stress As Modulators of Gut Microbiota - 2020 - NeurobiologySabrina YenNo ratings yet
- The Gut Brain-Axis in Neurological Diseases: Review ArticleDocument9 pagesThe Gut Brain-Axis in Neurological Diseases: Review ArticleDanielle GraceNo ratings yet
- From Probiotics To Psychobiotics Live BeneficialDocument22 pagesFrom Probiotics To Psychobiotics Live BeneficialDaniele GomesNo ratings yet
- Can Microbiology Affect Psychiatry A LinDocument17 pagesCan Microbiology Affect Psychiatry A LinSimona BengheNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument24 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsEcaterina Borovic-PavlovschiNo ratings yet
- Pharmaceutical NanotechnologyDocument39 pagesPharmaceutical Nanotechnologysaniyakulsum235No ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument19 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsCHIRANJEEVINo ratings yet
- 1 s2.0 S2090123221001867 MainDocument22 pages1 s2.0 S2090123221001867 MainkamelmafuNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument19 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For Scientistscalica laksmana100% (1)
- On The Organization of A Drug Discovery PlatformDocument39 pagesOn The Organization of A Drug Discovery PlatformBadou NindoNo ratings yet
- 47 - An Update On The Relationship Between The Gut Microbiome and Obsessive-Compulsive DisorderDocument11 pages47 - An Update On The Relationship Between The Gut Microbiome and Obsessive-Compulsive DisorderPaula CantalapiedraNo ratings yet
- Gastrointestinal Microbiology - A. Ouwehand, E. Vaughan (Taylor and Francis, 2006) WW PDFDocument431 pagesGastrointestinal Microbiology - A. Ouwehand, E. Vaughan (Taylor and Francis, 2006) WW PDFGabriela Balan100% (2)
- Metabonomics and Gut Microbiota in Nutrition and Disease: Sunil Kochhar François-Pierre Martin EditorsDocument389 pagesMetabonomics and Gut Microbiota in Nutrition and Disease: Sunil Kochhar François-Pierre Martin Editorserika100% (1)
- AntioxidantsDocument14 pagesAntioxidantsReena SapruNo ratings yet
- The Effect of Bacteria On DepressionDocument2 pagesThe Effect of Bacteria On DepressionLevi AckermanNo ratings yet
- Bryophytes A Potential Source of AntioxidantsDocument13 pagesBryophytes A Potential Source of AntioxidantsROBERTO MIRANDA CALDASNo ratings yet
- Scitranslmed 3000322Document12 pagesScitranslmed 3000322Arizha IrmaNo ratings yet
- Fundamentals of Microbiome Science: How Microbes Shape Animal BiologyFrom EverandFundamentals of Microbiome Science: How Microbes Shape Animal BiologyRating: 3 out of 5 stars3/5 (2)
- New GAP Algorithm Draft 4-7-19Document1 pageNew GAP Algorithm Draft 4-7-19Umber AgarwalNo ratings yet
- Package Contents Instruction Installation / Technical SupportDocument1 pagePackage Contents Instruction Installation / Technical SupportXseed02No ratings yet
- Braille PDFDocument6 pagesBraille PDFSarah SalamatNo ratings yet
- Love Languages Mystery GameDocument3 pagesLove Languages Mystery GamePittipat Manzano ChupungcoNo ratings yet
- 28 ESL Discussion TopicsDocument21 pages28 ESL Discussion TopicsMaruthi Maddipatla50% (2)
- Butterfly Lesson PlanDocument2 pagesButterfly Lesson Planapi-272455011No ratings yet
- Evaluating WebsitesDocument2 pagesEvaluating Websitesapi-322361374No ratings yet
- Final Exam Form 1Document12 pagesFinal Exam Form 1ILANGGO KANANNo ratings yet
- 8th Math EM TB 2022Document288 pages8th Math EM TB 2022Ramesh BabuNo ratings yet
- Arts. 1226 1230Document13 pagesArts. 1226 1230Vj McNo ratings yet
- Daily Loyalty Report 23-02-2022Document119 pagesDaily Loyalty Report 23-02-2022Jos ButtlerNo ratings yet
- RWD 3 SpeedDocument6 pagesRWD 3 SpeedAbraham VegaNo ratings yet
- Position Paper On DivorceDocument4 pagesPosition Paper On Divorcejereka100% (2)
- Chemical Engineering Department, Faculty of Engineering, Universitas Indonesia, Depok, West Java, IndonesiaDocument6 pagesChemical Engineering Department, Faculty of Engineering, Universitas Indonesia, Depok, West Java, IndonesiaBadzlinaKhairunizzahraNo ratings yet
- CH 2Document28 pagesCH 2younis ahmadNo ratings yet
- BehaviorismDocument21 pagesBehaviorismIqbal Haziq100% (2)
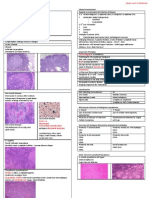
- Lymph Node PathologyDocument4 pagesLymph Node PathologyGerardLum0% (1)
- Peckiana Peckiana Peckiana Peckiana PeckianaDocument29 pagesPeckiana Peckiana Peckiana Peckiana PeckianaWais Al-QorniNo ratings yet
- A Glossary of The Words and Phrases of Furness North Lancashire 1000216573Document272 pagesA Glossary of The Words and Phrases of Furness North Lancashire 1000216573Mohammad WaseemNo ratings yet
- Outrageous Mantra ExperimentDocument22 pagesOutrageous Mantra ExperimentPopovici DraganNo ratings yet
- Rajni Vashisht Ashwani TambatDocument10 pagesRajni Vashisht Ashwani Tambatvaibhav bansalNo ratings yet
- ContingentWorker OffboardingTicketDocument7 pagesContingentWorker OffboardingTicketSonia Cuenca AcaroNo ratings yet
- 12 Siochi Vs GozonDocument10 pages12 Siochi Vs GozonPaolo CruzNo ratings yet
- Why Does AFRICA Called The Dark ContinentDocument11 pagesWhy Does AFRICA Called The Dark ContinentGio LagadiaNo ratings yet
- Eldon Riggs Hugie, Esq. Honored As A Professional of The Year For Five Consecutive Years by Strathmore's Who's Who Worldwide PublicationDocument2 pagesEldon Riggs Hugie, Esq. Honored As A Professional of The Year For Five Consecutive Years by Strathmore's Who's Who Worldwide PublicationPR.comNo ratings yet
- Footing Design Calculations: DefinitionsDocument31 pagesFooting Design Calculations: DefinitionsMark Roger Huberit IINo ratings yet
- Case Session - 8Document4 pagesCase Session - 8Shinas AhmmedNo ratings yet