Download as odt, pdf, or txt
You might also like
- DR - Amy Yasko Microbes Metals MethylationDocument211 pagesDR - Amy Yasko Microbes Metals Methylationmaimutalin100% (2)
- Anatomy and Physiology of ZombiesDocument12 pagesAnatomy and Physiology of ZombiesdsolisNo ratings yet
- Food Microbiology: Zhou Jianzhong, Liu Xiaoli, Jiang Hanhu, Dong MingshengDocument6 pagesFood Microbiology: Zhou Jianzhong, Liu Xiaoli, Jiang Hanhu, Dong MingshengRodolfo ChabléNo ratings yet
- Prevalence and Characterization of Cronobacter Spp. From Various Foods, Medicinal Plants, and Environmental SamplesDocument8 pagesPrevalence and Characterization of Cronobacter Spp. From Various Foods, Medicinal Plants, and Environmental Samplesjuniper leeNo ratings yet
- EztnDocument10 pagesEztnplastioid4079No ratings yet
- Expression of Flak Flagellin From Salmonella Typhimurium in Tobacco SeedsDocument4 pagesExpression of Flak Flagellin From Salmonella Typhimurium in Tobacco SeedsIOSR Journal of PharmacyNo ratings yet
- 511 jmm000460Document6 pages511 jmm000460Nur LaylaNo ratings yet
- Primer DesDocument6 pagesPrimer DesAhmed almahdiNo ratings yet
- B Lactam ChipDocument6 pagesB Lactam ChipBd HanNo ratings yet
- Analytical Biochemistry: Claudia Lefimil, Carla Lozano, Irene Morales-Bozo, Anita Plaza, Cristian Maturana, Blanca UrzúaDocument3 pagesAnalytical Biochemistry: Claudia Lefimil, Carla Lozano, Irene Morales-Bozo, Anita Plaza, Cristian Maturana, Blanca UrzúaWill BustNo ratings yet
- Rambabu NarvaneniDocument4 pagesRambabu NarvaneniS291991No ratings yet
- Detection and Typing of Human Papillomaviruses by PCRDocument5 pagesDetection and Typing of Human Papillomaviruses by PCRMyat OoNo ratings yet
- Golkar and JamilDocument6 pagesGolkar and JamilZhabiz GNo ratings yet
- Shatalin SomDocument25 pagesShatalin Somreinafeng1No ratings yet
- Science - Adh7950 SMDocument44 pagesScience - Adh7950 SMwhoselaundryNo ratings yet
- 10.1002@1097 0290 (20001020) 70 2 217 Aid Bit11 3.0.co 2 WDocument8 pages10.1002@1097 0290 (20001020) 70 2 217 Aid Bit11 3.0.co 2 WVenkata Suryanarayana GorleNo ratings yet
- Aem 02081-12Document5 pagesAem 02081-12Dylan TeránNo ratings yet
- The Veterinary Journal: Jun Zhang, Gui-Hong Zhang, Lin Yang, Ren Huang, Yu Zhang, Kun Jia, Wen Yuan, Shou-Jun LiDocument4 pagesThe Veterinary Journal: Jun Zhang, Gui-Hong Zhang, Lin Yang, Ren Huang, Yu Zhang, Kun Jia, Wen Yuan, Shou-Jun LiEdgar DíazNo ratings yet
- CPG Island Hypermethylation-Associated Silencing of Micrornas Promotes Human Endometrial CancerDocument10 pagesCPG Island Hypermethylation-Associated Silencing of Micrornas Promotes Human Endometrial CancerFerdina NidyasariNo ratings yet
- Advances in Biomolecular Medicine - Molecular Detection of DHA-1 AmpC Beta-Lactamase Gene in Enterobacteriaceae Clinical Isolates in IndonesiaDocument5 pagesAdvances in Biomolecular Medicine - Molecular Detection of DHA-1 AmpC Beta-Lactamase Gene in Enterobacteriaceae Clinical Isolates in IndonesiabungaNo ratings yet
- Enhanced Foreign Protein Accumulation in Nicotiana Benthamiana Leaves Co-Infiltrated With A TMV Vector and Plant Cell Cycle Regulator GenesDocument7 pagesEnhanced Foreign Protein Accumulation in Nicotiana Benthamiana Leaves Co-Infiltrated With A TMV Vector and Plant Cell Cycle Regulator Geneszhaoyue12112001No ratings yet
- Development of Polymerase Chain Reaction Assays With Host-Specific Internal Controls For Chlamydophila AbortusDocument7 pagesDevelopment of Polymerase Chain Reaction Assays With Host-Specific Internal Controls For Chlamydophila Abortusangela nandaNo ratings yet
- 1-Eastern Mediterranean Health JournalDocument5 pages1-Eastern Mediterranean Health Journalreadyboy89No ratings yet
- 2 Klebsiella PneumoniaeDocument11 pages2 Klebsiella PneumoniaeGhadah AlyousifNo ratings yet
- Genetic Diversity of The Prolactin Gene in Three Indonesian DucksDocument5 pagesGenetic Diversity of The Prolactin Gene in Three Indonesian DucksSanjaya RahmatNo ratings yet
- Sortase A Confers Protection Against Streptococcus Pneumoniae in MiceDocument5 pagesSortase A Confers Protection Against Streptococcus Pneumoniae in MiceRin ChanNo ratings yet
- Jurnal Sismik 1Document3 pagesJurnal Sismik 1Surasta MitaNo ratings yet
- Improving the production of transgenic fish germlines: In vivo evaluation of mosaicism in zebrafish (Danio rerio) using a green fluorescent protein (GFP) and growth hormone cDNA transgene co-injection strategyDocument6 pagesImproving the production of transgenic fish germlines: In vivo evaluation of mosaicism in zebrafish (Danio rerio) using a green fluorescent protein (GFP) and growth hormone cDNA transgene co-injection strategyEndah Permata SariNo ratings yet
- A. NigerDocument5 pagesA. NigerTran Van HoangNo ratings yet
- Cridia SergipensisDocument19 pagesCridia SergipensisedugersNo ratings yet
- Expression of Gene From A Novel Strain SY49-1 Active On Pest InsectsDocument6 pagesExpression of Gene From A Novel Strain SY49-1 Active On Pest InsectsYounglife UGNo ratings yet
- BAM CronobacterDocument15 pagesBAM CronobacterLACEN - Divisao de ProdutosNo ratings yet
- Isolation, Characterization and Expression of A Human Brain Mitochondrial Glutaminase cDNADocument8 pagesIsolation, Characterization and Expression of A Human Brain Mitochondrial Glutaminase cDNAMadhuri PatelNo ratings yet
- Molecular Characterization of ESBL-producing Shigella Sonnei Isolates From Patients With Bacilliary Dysentery in LebanonDocument6 pagesMolecular Characterization of ESBL-producing Shigella Sonnei Isolates From Patients With Bacilliary Dysentery in Lebanonvn_ny84bio021666No ratings yet
- 2013, Shigemori Et Al PDFDocument6 pages2013, Shigemori Et Al PDFDavid CampuésNo ratings yet
- Campylobacter Jejuni, C. Coli, C. Lari, C. UpsaliensisDocument4 pagesCampylobacter Jejuni, C. Coli, C. Lari, C. Upsaliensismalwylie100% (1)
- HLG PaperDocument6 pagesHLG PaperayanNo ratings yet
- Jmbfs 875 CazaDocument4 pagesJmbfs 875 CazaDorianNo ratings yet
- Eustaqio 2005 AntibioticsDocument8 pagesEustaqio 2005 AntibioticsXimena CortesNo ratings yet
- PALMEIRA Et Al, 2018Document2 pagesPALMEIRA Et Al, 2018MARIA FERNANDA OLIVEIRA SOARESNo ratings yet
- Development of An Allele-Specific, Loop-Mediated, Isothermal Amplification Method (AS-LAMP) To Detect The L1014F KDR-W Mutation in Anopheles Gambiae S. LDocument7 pagesDevelopment of An Allele-Specific, Loop-Mediated, Isothermal Amplification Method (AS-LAMP) To Detect The L1014F KDR-W Mutation in Anopheles Gambiae S. Librahima1968No ratings yet
- Antimicrobial Peptides in Oyster Hemolymph - The Bacterial Connection Version HALDocument48 pagesAntimicrobial Peptides in Oyster Hemolymph - The Bacterial Connection Version HALAmadorRevillaNo ratings yet
- 2003 Common Surface-Antigen Var Genes of Limited Diversity Expressed by Plasmodium Falciparum Placental Isolates Separated by Time and SpaceDocument8 pages2003 Common Surface-Antigen Var Genes of Limited Diversity Expressed by Plasmodium Falciparum Placental Isolates Separated by Time and SpaceSethawud ChaikitgosiyakulNo ratings yet
- PrimerDocument5 pagesPrimerrfsh rfshNo ratings yet
- Molecular Detection of Resistance and Virulence Genes in Coagulase Negative Staphylococci Isolated From Blood Cultures at The University Teaching Hospital of BouakeDocument11 pagesMolecular Detection of Resistance and Virulence Genes in Coagulase Negative Staphylococci Isolated From Blood Cultures at The University Teaching Hospital of BouakeSambhavNo ratings yet
- Weisburg Et Al 1991 16s Ribosomal Dna Amplification For Phylogenetic StudyDocument7 pagesWeisburg Et Al 1991 16s Ribosomal Dna Amplification For Phylogenetic Studyteacher.mireyaNo ratings yet
- Comparative DNA Nalysis of Oconut (A C Cocos Nucifera L.) Palms With Polyembryonic and Monoembryonic OriginsDocument7 pagesComparative DNA Nalysis of Oconut (A C Cocos Nucifera L.) Palms With Polyembryonic and Monoembryonic OriginsVanessa S. MacaleleNo ratings yet
- 9140-Article Text-17119-1-10-20200716Document8 pages9140-Article Text-17119-1-10-20200716Maha hameedNo ratings yet
- 10.33073 - PJM 2022 023Document6 pages10.33073 - PJM 2022 023Andy SetiawanNo ratings yet
- Molecular Characterization of The Isolated Strains of Bovine Viral Diarrhea VirusDocument7 pagesMolecular Characterization of The Isolated Strains of Bovine Viral Diarrhea VirusGhada AfifiNo ratings yet
- Myeloid NeoplasiaDocument10 pagesMyeloid NeoplasiaivssonNo ratings yet
- Ensayos de Alto Rendimiento para La Detección de La Mutación F1534C en El Gen Del Canal de Sodio Dependiente de VoltajeDocument9 pagesEnsayos de Alto Rendimiento para La Detección de La Mutación F1534C en El Gen Del Canal de Sodio Dependiente de VoltajeJuan Miguel Castro CamachoNo ratings yet
- Detection of Toxoplasma Gondii by PCR and Quantitative PCR With High Specificty and Lower Limit of DetectionDocument7 pagesDetection of Toxoplasma Gondii by PCR and Quantitative PCR With High Specificty and Lower Limit of DetectionSholatya FurqonieNo ratings yet
- Weisburg Et Al 1991 16s Ribosomal Dna Amplification For Phylogenetic StudyDocument7 pagesWeisburg Et Al 1991 16s Ribosomal Dna Amplification For Phylogenetic Studyteacher.mireyaNo ratings yet
- Comparison Lamp Con OtrasDocument7 pagesComparison Lamp Con OtrasLuis Ernesto Mena ZapataNo ratings yet
- JM 002076Document5 pagesJM 002076Malú CarranzaNo ratings yet
- www_vet-202110-0003Document9 pageswww_vet-202110-0003minhclu333No ratings yet
- General Biology 2: Genetic EngineeringDocument11 pagesGeneral Biology 2: Genetic EngineeringHazel AbitriaNo ratings yet
- The Anaplasma Marginale msp5 Gene Encodes A 19 - Kilodalton Protein Conserved in All Recognized Anaplasma SpeciesDocument6 pagesThe Anaplasma Marginale msp5 Gene Encodes A 19 - Kilodalton Protein Conserved in All Recognized Anaplasma Speciesyudhi arjentiniaNo ratings yet
- Epidermidis Carrying Biofilm Formation GenesDocument5 pagesEpidermidis Carrying Biofilm Formation GenesLini MaliqisnayantiNo ratings yet
- Medical Mycology: Cellular and Molecular TechniquesFrom EverandMedical Mycology: Cellular and Molecular TechniquesKevin KavanaghNo ratings yet
- DNS Bhuli Timetable Portion (Online Test)Document20 pagesDNS Bhuli Timetable Portion (Online Test)Himanshu RajNo ratings yet
- Chapter 2 Biodiversity 生物多样性: Diversity 多样化 of Organisms 生物Document138 pagesChapter 2 Biodiversity 生物多样性: Diversity 多样化 of Organisms 生物YAP SHAN MoeNo ratings yet
- Amino Acids, Proteins, and EnzymesDocument62 pagesAmino Acids, Proteins, and Enzymesendale gebregzabherNo ratings yet
- Tietz Textbook of Clinical Chemistry and Mo... : by Carl A. Burtis PHDDocument13 pagesTietz Textbook of Clinical Chemistry and Mo... : by Carl A. Burtis PHDReg LagartejaNo ratings yet
- (Methods in Molecular Biology 1189) Celeste M. Nelson (Eds.) - Tissue Morphogenesis - Methods and Protocols-Humana Press (2015)Document365 pages(Methods in Molecular Biology 1189) Celeste M. Nelson (Eds.) - Tissue Morphogenesis - Methods and Protocols-Humana Press (2015)Pranav Vyas100% (1)
- Student Project: Applications of Exponential Functions: Pure Mathematics 30Document8 pagesStudent Project: Applications of Exponential Functions: Pure Mathematics 30Pie PeyNo ratings yet
- Characteristics of FungiDocument1 pageCharacteristics of FungiPaige DarbonneNo ratings yet
- Anatomy and Physiology ResourcesDocument4 pagesAnatomy and Physiology Resourcesmasmakkin6170No ratings yet
- GENBIO2 - Lesson - The Central Dogma of Molecular BiologyDocument2 pagesGENBIO2 - Lesson - The Central Dogma of Molecular BiologyJazmaine SimbulanNo ratings yet
- Stiles-Jernigan - The Basics of Brain DevelopmentDocument22 pagesStiles-Jernigan - The Basics of Brain DevelopmentBrunela SperanzaNo ratings yet
- Immunoparasitology and Fungal ImmunityDocument31 pagesImmunoparasitology and Fungal ImmunityShakti PatelNo ratings yet
- Molecular Clock Dating Using Mrbayes: Chi Zhang May 28, 2018Document25 pagesMolecular Clock Dating Using Mrbayes: Chi Zhang May 28, 2018John Marie FamosoNo ratings yet
- Activity 4. Culture Media PreparationDocument4 pagesActivity 4. Culture Media PreparationRoan Eam TanNo ratings yet
- Obat Herbal Untuk AritmiaDocument17 pagesObat Herbal Untuk AritmiaFerina Nadya PratamaNo ratings yet
- Food Microbiology William Carroll FrazierDocument490 pagesFood Microbiology William Carroll FrazierK V SATHEESHKUMAR MECHNo ratings yet
- Pollination: and Its TypesDocument15 pagesPollination: and Its TypessiddNo ratings yet
- Mechanotransduction PDFDocument496 pagesMechanotransduction PDFvNo ratings yet
- Genera of Phytopathogenic Fungi GOPHY3Document124 pagesGenera of Phytopathogenic Fungi GOPHY3WilliamNo ratings yet
- Meiosis Intro Haploid and DiploidDocument48 pagesMeiosis Intro Haploid and DiploidJudy RianoNo ratings yet
- The Emerging Field of Human Social GenomicsDocument18 pagesThe Emerging Field of Human Social GenomicsEdilson RodriguesNo ratings yet
- PDF The Vegetable Pathosystem Ecology Disease Mechanism and Management 1St Edition Mohammad Ansar Editor Ebook Full ChapterDocument53 pagesPDF The Vegetable Pathosystem Ecology Disease Mechanism and Management 1St Edition Mohammad Ansar Editor Ebook Full Chapterjackie.robinson980100% (1)
- Microbiology & ImmunologyDocument3 pagesMicrobiology & ImmunologycknsyNo ratings yet
- Tu BSC Microbiology 1st Year SyllabusDocument71 pagesTu BSC Microbiology 1st Year SyllabusSujan PaudelNo ratings yet
- Pengertian Dan Sejarah Bioteknologi - 2021Document39 pagesPengertian Dan Sejarah Bioteknologi - 2021Nila FirmaliaNo ratings yet
- Dot Blot ProtocolDocument2 pagesDot Blot ProtocolSuki BlytheNo ratings yet
- Assignment For The Fourth Semesters Botany Department ST MarysDocument3 pagesAssignment For The Fourth Semesters Botany Department ST MarysRimeiaNo ratings yet
- Kingdom FungiDocument11 pagesKingdom FungiMihnea RotaruNo ratings yet
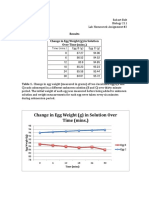
- Results Change in Egg Weight (G) in Solution Over Time (Mins.)Document2 pagesResults Change in Egg Weight (G) in Solution Over Time (Mins.)Roberto BeltNo ratings yet