
PIIS0021925818434217
PIIS0021925818434217
You might also like
- Royal FamilyDocument18 pagesRoyal FamilyMarrauNo ratings yet
- Niacinamide Salicylate ComplexDocument4 pagesNiacinamide Salicylate ComplexMarrauNo ratings yet
- KOHJIN Glutathione Bioavailability (ACS)Document7 pagesKOHJIN Glutathione Bioavailability (ACS)806328366No ratings yet
- The Effect of The Sodium-Glucose Cotransporter Type-2 Inhibitor Dapagliflozin On Glomerular Filtration Rate in Healthy CatsDocument6 pagesThe Effect of The Sodium-Glucose Cotransporter Type-2 Inhibitor Dapagliflozin On Glomerular Filtration Rate in Healthy CatsJohn QuickNo ratings yet
- Bagus GlomerulosclerosisDocument10 pagesBagus GlomerulosclerosisJundiNo ratings yet
- Jcinvest00134 0257Document13 pagesJcinvest00134 0257Eva RefitaNo ratings yet
- Zhang., 2003. Mekanisme a`-Go-Go-related Gene (HERG) Pada Pasien DM Terhadap Prolong QTDocument11 pagesZhang., 2003. Mekanisme a`-Go-Go-related Gene (HERG) Pada Pasien DM Terhadap Prolong QThasan andrianNo ratings yet
- Blood 6Document13 pagesBlood 6ashokNo ratings yet
- Clin Chim Acta. 1998 - 275 - 175-184 Stefano 3Document10 pagesClin Chim Acta. 1998 - 275 - 175-184 Stefano 3scopettiNo ratings yet
- Age Related Changes in Glutathione Concentration Glutathione Peroxidase Glutathione S Transferase and Superoxide Dismutase Activities in SenescenceDocument5 pagesAge Related Changes in Glutathione Concentration Glutathione Peroxidase Glutathione S Transferase and Superoxide Dismutase Activities in SenescenceTron TronNo ratings yet
- The Relationship Between The Level of Glutathione, Impairment of Glucose Metabolism and Complications of Diabetes MellitusDocument5 pagesThe Relationship Between The Level of Glutathione, Impairment of Glucose Metabolism and Complications of Diabetes MellitusKikiaraNo ratings yet
- Diabetes 2001 RonnerDocument10 pagesDiabetes 2001 RonnerGerardo Félix MartínezNo ratings yet
- Enzymic Method For Quantitative Determination of Nanogram Amounts of Total and Oxidized GlutathioneDocument21 pagesEnzymic Method For Quantitative Determination of Nanogram Amounts of Total and Oxidized GlutathioneJosué Velázquez100% (1)
- (14796805 - Journal of Endocrinology) Glutamate Dehydrogenase, Insulin Secretion, And Type 2 Diabetes - a New Means to Protect the Pancreatic Β-cellDocument4 pages(14796805 - Journal of Endocrinology) Glutamate Dehydrogenase, Insulin Secretion, And Type 2 Diabetes - a New Means to Protect the Pancreatic Β-cellVelani ElendiaNo ratings yet
- BR J Haematol - 2007 - Vasavda - The Linear Effects of Thalassaemia The UGT1A1 and HMOX1 Polymorphisms On CholelithiasisDocument8 pagesBR J Haematol - 2007 - Vasavda - The Linear Effects of Thalassaemia The UGT1A1 and HMOX1 Polymorphisms On CholelithiasisJordan NGOUNENo ratings yet
- 8-Adrenergic Stimulatory GTP-binding Protein of Adenylate: Functional Reconstitution of Receptors and The CyclaseDocument5 pages8-Adrenergic Stimulatory GTP-binding Protein of Adenylate: Functional Reconstitution of Receptors and The CyclaseSean Christopher PawlowskiNo ratings yet
- One Systemic Protect Ant Against Oxidative and Free Radical DamageDocument19 pagesOne Systemic Protect Ant Against Oxidative and Free Radical DamageNicky ElsworthNo ratings yet
- Role of N Acetylcysteine and Cystine in Glutathione Synthesis in Human ErythrocytesDocument11 pagesRole of N Acetylcysteine and Cystine in Glutathione Synthesis in Human ErythrocytesVolkan YprkcNo ratings yet
- BIOchem - Glucose - Tolerance - Report - ) TOMDocument20 pagesBIOchem - Glucose - Tolerance - Report - ) TOMmujuni emanuelNo ratings yet
- Decreased Ligand Affinity Rather Than Glucocorticoid Receptor Down-Regulation in Patients With Endogenous Cushing's SyndromeDocument5 pagesDecreased Ligand Affinity Rather Than Glucocorticoid Receptor Down-Regulation in Patients With Endogenous Cushing's SyndromeParishan SaeedNo ratings yet
- The Effects of Hyperglycemia and Endotoxemia On Coagulation Parameters inDocument7 pagesThe Effects of Hyperglycemia and Endotoxemia On Coagulation Parameters inGrasiene MenesesNo ratings yet
- The Effects of Green Teaaaa 2015Document12 pagesThe Effects of Green Teaaaa 2015Darius HădăreanNo ratings yet
- Protein Kinase Cis Activated in From Rats: Glomeruli DiabeticDocument9 pagesProtein Kinase Cis Activated in From Rats: Glomeruli Diabeticapi-19757116No ratings yet
- KOHJIN Glutathione Absorption MechanismDocument8 pagesKOHJIN Glutathione Absorption Mechanism806328366No ratings yet
- J. Biol. Chem.-1988-Meister-17205-8Document4 pagesJ. Biol. Chem.-1988-Meister-17205-8Pepé DubNo ratings yet
- Chemlab 9 CarbohydratesDocument8 pagesChemlab 9 CarbohydratesirfanNo ratings yet
- TMP 83 BBDocument10 pagesTMP 83 BBFrontiersNo ratings yet
- J. Bacteriol.-2010-Daddaoua-4357-66 PDFDocument11 pagesJ. Bacteriol.-2010-Daddaoua-4357-66 PDFAmelia PratiwiNo ratings yet
- Alpha Lactalbumin To HamletDocument6 pagesAlpha Lactalbumin To Hamlet6478505032No ratings yet
- 1 s2.0 S2211383513000129 Main PDFDocument5 pages1 s2.0 S2211383513000129 Main PDFPili CárdenasNo ratings yet
- Articulo Biologia CelularDocument8 pagesArticulo Biologia CelularLeonardo SánchezNo ratings yet
- 0 GlucagonDocument12 pages0 GlucagonRenata Lima de FreitasNo ratings yet
- IsotachophoresisDocument7 pagesIsotachophoresisEka HerlinaNo ratings yet
- ISSN: 1756-2228: Home Editors Authors Ambassadors Contact News SearchDocument36 pagesISSN: 1756-2228: Home Editors Authors Ambassadors Contact News SearchanakfkubNo ratings yet
- p1541 - Vol 84 - 9 PDFDocument5 pagesp1541 - Vol 84 - 9 PDFmresearchNo ratings yet
- B3 Niacin Energy MitochondriaDocument15 pagesB3 Niacin Energy MitochondriavalgurNo ratings yet
- Empaglifozin, Vía Switching Metabolism Towards Lipid Uitlization, Moderately Increase LDL-Cholesterol Level Through Reduced LDL CatabolimsDocument20 pagesEmpaglifozin, Vía Switching Metabolism Towards Lipid Uitlization, Moderately Increase LDL-Cholesterol Level Through Reduced LDL CatabolimsLaura CalderónNo ratings yet
- International Journal of Pharmaceutical and Clinical ScienceDocument3 pagesInternational Journal of Pharmaceutical and Clinical ScienceJamesNo ratings yet
- Antidiabetic Effects of Quercetin in Streptozocin-Induced Diabetic RatsDocument8 pagesAntidiabetic Effects of Quercetin in Streptozocin-Induced Diabetic RatsMohammad IvanNo ratings yet
- Vivo Analyses of Experimental Animals (6,7)Document23 pagesVivo Analyses of Experimental Animals (6,7)aakash sahaNo ratings yet
- 10.1515 - Labmed 2019 0098Document9 pages10.1515 - Labmed 2019 0098Fery ArdiansahNo ratings yet
- Cell Biochem Funct 2011 p55Document9 pagesCell Biochem Funct 2011 p55IQBAL HAKKIKINo ratings yet
- Extraction of Glycogen On Mild Condition Lacks Aig FractionDocument7 pagesExtraction of Glycogen On Mild Condition Lacks Aig FractionMaika DoronelaNo ratings yet
- Hyperglycemia As An Effect of Cardiopulmonary Bypass: Intra-Operative Glucose ManagementDocument6 pagesHyperglycemia As An Effect of Cardiopulmonary Bypass: Intra-Operative Glucose ManagementMuhammad RizqiNo ratings yet
- Liraglutide Improves Lipid Metabolism by Enhancing Cholesterol Efflux Associated With ABCA1 and ERK1/2 PathwayDocument12 pagesLiraglutide Improves Lipid Metabolism by Enhancing Cholesterol Efflux Associated With ABCA1 and ERK1/2 PathwayAndrianna NastasyaNo ratings yet
- Reti ManDocument8 pagesReti ManSUMIT PANDEYNo ratings yet
- Renal Threshold For Glucose ReabsorptionDocument4 pagesRenal Threshold For Glucose ReabsorptionsoniNo ratings yet
- Free Radical Biology and Medicine: Invited Review ArticleDocument13 pagesFree Radical Biology and Medicine: Invited Review ArticleNatalia SimõesNo ratings yet
- Re Glucose Reprt-1Document23 pagesRe Glucose Reprt-1Ingrid BayiyanaNo ratings yet
- Cancer Cell Get Fat From Glutamine 2012Document4 pagesCancer Cell Get Fat From Glutamine 2012Yulianti RiskiNo ratings yet
- ProImmune F1 On Adenine-Induced Chronic Progressive Tubulointerstitial NephropathyDocument12 pagesProImmune F1 On Adenine-Induced Chronic Progressive Tubulointerstitial NephropathyBryan Edson M. YcoNo ratings yet
- Methods: A Comparison of Three of Glycogen Measurement in TissuesDocument8 pagesMethods: A Comparison of Three of Glycogen Measurement in TissuesDawn KentNo ratings yet
- Ijms 21 07833Document25 pagesIjms 21 07833Zerrae CBTNo ratings yet
- Pradedova 2018Document9 pagesPradedova 2018rocio fernandez san juanNo ratings yet
- Glutathione Antioxidant Properties Dedicated To NaDocument21 pagesGlutathione Antioxidant Properties Dedicated To NaTazkiyatul Asma'iNo ratings yet
- A Novel Therapeutic Agent For Type 2 Diabetes Mellitus: SGLT2 InhibitorDocument13 pagesA Novel Therapeutic Agent For Type 2 Diabetes Mellitus: SGLT2 InhibitorSheldon SilvaNo ratings yet
- Sun, 2010 DHA Regulates Lipogenesis and Lipolysis Genes in Mice Adipose and LiverDocument7 pagesSun, 2010 DHA Regulates Lipogenesis and Lipolysis Genes in Mice Adipose and LiverDaniela Patricia Alvarez AravenaNo ratings yet
- HHS Public Access: Sodium-Glucose CotransportDocument12 pagesHHS Public Access: Sodium-Glucose CotransportAlexandra VásquezNo ratings yet
- MJMS30022023 09Document15 pagesMJMS30022023 09Muzammal RazaNo ratings yet
- CDD 2011102 ADocument12 pagesCDD 2011102 ACaerulus Fuad Abdul BaqiNo ratings yet
- 2.glycolysis & Oxidation of PyruvateDocument12 pages2.glycolysis & Oxidation of Pyruvateقتيبه خالد دحام خلفNo ratings yet
- Rompf Icfp15Document8 pagesRompf Icfp15MarrauNo ratings yet
- Tetrodotoxin: TTX: BackgroundDocument12 pagesTetrodotoxin: TTX: BackgroundMarrauNo ratings yet
- Royal FamilyDocument38 pagesRoyal FamilyMarrauNo ratings yet
- 2:4:6-Trinitrotoluene: 0-0114N-Ba (OH)Document16 pages2:4:6-Trinitrotoluene: 0-0114N-Ba (OH)MarrauNo ratings yet
- Penerapan Prinsip Rule of Reason Pada Putusan Perkara Nomor 08/KPPU-I/2020 Tentang Dugaan Praktik Diskriminasi Antara Telkom-Telkomsel Dan NetflixDocument14 pagesPenerapan Prinsip Rule of Reason Pada Putusan Perkara Nomor 08/KPPU-I/2020 Tentang Dugaan Praktik Diskriminasi Antara Telkom-Telkomsel Dan NetflixMarrauNo ratings yet
- Examining Explosives Handling Sensitivity of Trinitrotoluene (TNT) With Different Particle SizesDocument7 pagesExamining Explosives Handling Sensitivity of Trinitrotoluene (TNT) With Different Particle SizesMarrauNo ratings yet
- Toxins 14 00576 v2Document34 pagesToxins 14 00576 v2MarrauNo ratings yet
- Molecules TNTDocument9 pagesMolecules TNTMarrauNo ratings yet
- Q. Exp.: A To NerveDocument10 pagesQ. Exp.: A To NerveMarrauNo ratings yet
- Effect Salicylic Acid To LettuceDocument6 pagesEffect Salicylic Acid To LettuceMarrauNo ratings yet
- Aspirin: Damocles Gruppe C Daniel Deckenbach Bilal Danisman Benedict Depp Scotty CobbDocument7 pagesAspirin: Damocles Gruppe C Daniel Deckenbach Bilal Danisman Benedict Depp Scotty CobbMarrauNo ratings yet
- Partial Characterization of Tetrodotoxin-Binding Component From Nerve MembraneDocument4 pagesPartial Characterization of Tetrodotoxin-Binding Component From Nerve MembraneMarrauNo ratings yet
- ICRI Sunscreen 2Document21 pagesICRI Sunscreen 2MarrauNo ratings yet
- The Synthesis, Properties, & Application of Salicylic Acid in The Treatment of AcneDocument13 pagesThe Synthesis, Properties, & Application of Salicylic Acid in The Treatment of AcneMarrauNo ratings yet
- Biosyntesis of RetinolDocument5 pagesBiosyntesis of RetinolMarrauNo ratings yet
- Treatment of PainDocument5 pagesTreatment of PainMarrauNo ratings yet
- Breast CancerDocument36 pagesBreast CancerMarrauNo ratings yet
- Vitamin B3 at A Glance: CancerDocument3 pagesVitamin B3 at A Glance: CancerMarrauNo ratings yet
- Nutritional Medicine and Diet Care: ClinmedDocument5 pagesNutritional Medicine and Diet Care: ClinmedMarrauNo ratings yet
- Salicylic AcidDocument5 pagesSalicylic AcidMarrauNo ratings yet
- Retinol Product InformationDocument3 pagesRetinol Product InformationMarrauNo ratings yet
- It First Into Retinoic Acid. Retinyl Palmitate: Layers Up, It Is Safe To Use With AhasDocument2 pagesIt First Into Retinoic Acid. Retinyl Palmitate: Layers Up, It Is Safe To Use With AhasMarrauNo ratings yet
- 2dot3 AgingSkin PhysRes1 BellemereGDocument10 pages2dot3 AgingSkin PhysRes1 BellemereGMarrauNo ratings yet
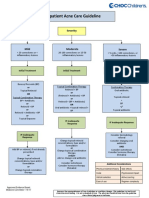
- Outpatient Acne Care Guideline: SeverityDocument5 pagesOutpatient Acne Care Guideline: SeverityMarrauNo ratings yet
- Public Law 113-195 113th Congress An ActDocument18 pagesPublic Law 113-195 113th Congress An ActMarrauNo ratings yet
- Retinol in CosmeticsDocument204 pagesRetinol in CosmeticsMarrauNo ratings yet
- Sunscreen Market AnalysisDocument43 pagesSunscreen Market AnalysisMarrauNo ratings yet
- En515 2017Document11 pagesEn515 2017katemizNo ratings yet
- Aromatic Reactions ExercisesDocument6 pagesAromatic Reactions Exercisesisaacrenzie.dipus.sciNo ratings yet
- 100 MTPD Acetic Acid Plant DesignDocument60 pages100 MTPD Acetic Acid Plant Designmudasserulbuet18No ratings yet
- MCQs of Atomic and Nuclear Physics With AnswersDocument8 pagesMCQs of Atomic and Nuclear Physics With Answers09whitedevil90No ratings yet
- Tech Sheet - ProSilver45TDocument2 pagesTech Sheet - ProSilver45TMicheal Booka WhelanNo ratings yet
- 9701_m24_qp_42Document28 pages9701_m24_qp_42rashya.sharmaNo ratings yet
- RIL J4 Projects 2Document25 pagesRIL J4 Projects 2sumit6singhNo ratings yet
- General ToxicologyDocument28 pagesGeneral Toxicologyhhes8116No ratings yet
- Natural PolymersDocument35 pagesNatural Polymerssiti sunarintyasNo ratings yet
- Safety Data Sheet of XI40211-000Document11 pagesSafety Data Sheet of XI40211-000irvingrscNo ratings yet
- ALTA 2020 NCC - Queensland Pacific MetalsDocument19 pagesALTA 2020 NCC - Queensland Pacific MetalssirbrianmiloNo ratings yet
- Agilent 5100 Series ICP-OES Consumables Ordering GuideDocument44 pagesAgilent 5100 Series ICP-OES Consumables Ordering Guideandri purnomoNo ratings yet
- Method 6.0 2014Document2 pagesMethod 6.0 2014FelisbertoNo ratings yet
- Chem 1008 Lab Midterms 2Document6 pagesChem 1008 Lab Midterms 2Jie Ann Faith AusmoloNo ratings yet
- Early Stage Recovery of Lithium From Tailored Thermal Conditioned Black Mass Part I (Schwich Et Al 2021)Document30 pagesEarly Stage Recovery of Lithium From Tailored Thermal Conditioned Black Mass Part I (Schwich Et Al 2021)irdantoNo ratings yet
- Practice Test-02 Arjuna NEET (2025) 09.06.2024 SolutionsDocument15 pagesPractice Test-02 Arjuna NEET (2025) 09.06.2024 Solutionskenyen.jarekNo ratings yet
- Ehudin Jacs ironIVoxoDocument19 pagesEhudin Jacs ironIVoxoneerajNo ratings yet
- MSDS MotorexDocument6 pagesMSDS MotorexGezy MegalittaNo ratings yet
- COA Soda Ash Dense Ansac - 230522 - 143613Document2 pagesCOA Soda Ash Dense Ansac - 230522 - 143613nur.hudaNo ratings yet
- Honeywell Genetron R410aDocument15 pagesHoneywell Genetron R410avivekNo ratings yet
- Yosim and MayerDocument66 pagesYosim and MayerRajabMumbeeNo ratings yet
- Ebook Chemistry For Today General Organic and Biochemistry PDF Full Chapter PDFDocument67 pagesEbook Chemistry For Today General Organic and Biochemistry PDF Full Chapter PDFrobert.davidson233100% (31)
- Ams 4911LDocument7 pagesAms 4911Ljamesdong1120No ratings yet
- Calculation Part 2 in AC-2Document2 pagesCalculation Part 2 in AC-223005852No ratings yet
- Grade 12 P2 2016 - 2023 papersDocument287 pagesGrade 12 P2 2016 - 2023 paperslihlemananga65No ratings yet
- Ebook Corrosion Atlas A Collection of Illustrated Case Histories PDF Full Chapter PDFDocument55 pagesEbook Corrosion Atlas A Collection of Illustrated Case Histories PDF Full Chapter PDFedwina.perry203100% (40)
- Module 1 (Thermo 2)Document38 pagesModule 1 (Thermo 2)Genesis MedelNo ratings yet
- KCET 2024 Chemistry Paper With AnswerDocument9 pagesKCET 2024 Chemistry Paper With Answerthejasmath2005No ratings yet
- CZ 132Document1 pageCZ 132Taner DemirelNo ratings yet
- Aldehydes, Ketones, and Carboxylic Acids Shobhit NirwanDocument12 pagesAldehydes, Ketones, and Carboxylic Acids Shobhit Nirwangokulss374100% (1)






















































































































Download as pdf or txt
You might also like
- Royal FamilyDocument18 pagesRoyal FamilyMarrauNo ratings yet
- Niacinamide Salicylate ComplexDocument4 pagesNiacinamide Salicylate ComplexMarrauNo ratings yet
- KOHJIN Glutathione Bioavailability (ACS)Document7 pagesKOHJIN Glutathione Bioavailability (ACS)806328366No ratings yet
- The Effect of The Sodium-Glucose Cotransporter Type-2 Inhibitor Dapagliflozin On Glomerular Filtration Rate in Healthy CatsDocument6 pagesThe Effect of The Sodium-Glucose Cotransporter Type-2 Inhibitor Dapagliflozin On Glomerular Filtration Rate in Healthy CatsJohn QuickNo ratings yet
- Bagus GlomerulosclerosisDocument10 pagesBagus GlomerulosclerosisJundiNo ratings yet
- Jcinvest00134 0257Document13 pagesJcinvest00134 0257Eva RefitaNo ratings yet
- Zhang., 2003. Mekanisme a`-Go-Go-related Gene (HERG) Pada Pasien DM Terhadap Prolong QTDocument11 pagesZhang., 2003. Mekanisme a`-Go-Go-related Gene (HERG) Pada Pasien DM Terhadap Prolong QThasan andrianNo ratings yet
- Blood 6Document13 pagesBlood 6ashokNo ratings yet
- Clin Chim Acta. 1998 - 275 - 175-184 Stefano 3Document10 pagesClin Chim Acta. 1998 - 275 - 175-184 Stefano 3scopettiNo ratings yet
- Age Related Changes in Glutathione Concentration Glutathione Peroxidase Glutathione S Transferase and Superoxide Dismutase Activities in SenescenceDocument5 pagesAge Related Changes in Glutathione Concentration Glutathione Peroxidase Glutathione S Transferase and Superoxide Dismutase Activities in SenescenceTron TronNo ratings yet
- The Relationship Between The Level of Glutathione, Impairment of Glucose Metabolism and Complications of Diabetes MellitusDocument5 pagesThe Relationship Between The Level of Glutathione, Impairment of Glucose Metabolism and Complications of Diabetes MellitusKikiaraNo ratings yet
- Diabetes 2001 RonnerDocument10 pagesDiabetes 2001 RonnerGerardo Félix MartínezNo ratings yet
- Enzymic Method For Quantitative Determination of Nanogram Amounts of Total and Oxidized GlutathioneDocument21 pagesEnzymic Method For Quantitative Determination of Nanogram Amounts of Total and Oxidized GlutathioneJosué Velázquez100% (1)
- (14796805 - Journal of Endocrinology) Glutamate Dehydrogenase, Insulin Secretion, And Type 2 Diabetes - a New Means to Protect the Pancreatic Β-cellDocument4 pages(14796805 - Journal of Endocrinology) Glutamate Dehydrogenase, Insulin Secretion, And Type 2 Diabetes - a New Means to Protect the Pancreatic Β-cellVelani ElendiaNo ratings yet
- BR J Haematol - 2007 - Vasavda - The Linear Effects of Thalassaemia The UGT1A1 and HMOX1 Polymorphisms On CholelithiasisDocument8 pagesBR J Haematol - 2007 - Vasavda - The Linear Effects of Thalassaemia The UGT1A1 and HMOX1 Polymorphisms On CholelithiasisJordan NGOUNENo ratings yet
- 8-Adrenergic Stimulatory GTP-binding Protein of Adenylate: Functional Reconstitution of Receptors and The CyclaseDocument5 pages8-Adrenergic Stimulatory GTP-binding Protein of Adenylate: Functional Reconstitution of Receptors and The CyclaseSean Christopher PawlowskiNo ratings yet
- One Systemic Protect Ant Against Oxidative and Free Radical DamageDocument19 pagesOne Systemic Protect Ant Against Oxidative and Free Radical DamageNicky ElsworthNo ratings yet
- Role of N Acetylcysteine and Cystine in Glutathione Synthesis in Human ErythrocytesDocument11 pagesRole of N Acetylcysteine and Cystine in Glutathione Synthesis in Human ErythrocytesVolkan YprkcNo ratings yet
- BIOchem - Glucose - Tolerance - Report - ) TOMDocument20 pagesBIOchem - Glucose - Tolerance - Report - ) TOMmujuni emanuelNo ratings yet
- Decreased Ligand Affinity Rather Than Glucocorticoid Receptor Down-Regulation in Patients With Endogenous Cushing's SyndromeDocument5 pagesDecreased Ligand Affinity Rather Than Glucocorticoid Receptor Down-Regulation in Patients With Endogenous Cushing's SyndromeParishan SaeedNo ratings yet
- The Effects of Hyperglycemia and Endotoxemia On Coagulation Parameters inDocument7 pagesThe Effects of Hyperglycemia and Endotoxemia On Coagulation Parameters inGrasiene MenesesNo ratings yet
- The Effects of Green Teaaaa 2015Document12 pagesThe Effects of Green Teaaaa 2015Darius HădăreanNo ratings yet
- Protein Kinase Cis Activated in From Rats: Glomeruli DiabeticDocument9 pagesProtein Kinase Cis Activated in From Rats: Glomeruli Diabeticapi-19757116No ratings yet
- KOHJIN Glutathione Absorption MechanismDocument8 pagesKOHJIN Glutathione Absorption Mechanism806328366No ratings yet
- J. Biol. Chem.-1988-Meister-17205-8Document4 pagesJ. Biol. Chem.-1988-Meister-17205-8Pepé DubNo ratings yet
- Chemlab 9 CarbohydratesDocument8 pagesChemlab 9 CarbohydratesirfanNo ratings yet
- TMP 83 BBDocument10 pagesTMP 83 BBFrontiersNo ratings yet
- J. Bacteriol.-2010-Daddaoua-4357-66 PDFDocument11 pagesJ. Bacteriol.-2010-Daddaoua-4357-66 PDFAmelia PratiwiNo ratings yet
- Alpha Lactalbumin To HamletDocument6 pagesAlpha Lactalbumin To Hamlet6478505032No ratings yet
- 1 s2.0 S2211383513000129 Main PDFDocument5 pages1 s2.0 S2211383513000129 Main PDFPili CárdenasNo ratings yet
- Articulo Biologia CelularDocument8 pagesArticulo Biologia CelularLeonardo SánchezNo ratings yet
- 0 GlucagonDocument12 pages0 GlucagonRenata Lima de FreitasNo ratings yet
- IsotachophoresisDocument7 pagesIsotachophoresisEka HerlinaNo ratings yet
- ISSN: 1756-2228: Home Editors Authors Ambassadors Contact News SearchDocument36 pagesISSN: 1756-2228: Home Editors Authors Ambassadors Contact News SearchanakfkubNo ratings yet
- p1541 - Vol 84 - 9 PDFDocument5 pagesp1541 - Vol 84 - 9 PDFmresearchNo ratings yet
- B3 Niacin Energy MitochondriaDocument15 pagesB3 Niacin Energy MitochondriavalgurNo ratings yet
- Empaglifozin, Vía Switching Metabolism Towards Lipid Uitlization, Moderately Increase LDL-Cholesterol Level Through Reduced LDL CatabolimsDocument20 pagesEmpaglifozin, Vía Switching Metabolism Towards Lipid Uitlization, Moderately Increase LDL-Cholesterol Level Through Reduced LDL CatabolimsLaura CalderónNo ratings yet
- International Journal of Pharmaceutical and Clinical ScienceDocument3 pagesInternational Journal of Pharmaceutical and Clinical ScienceJamesNo ratings yet
- Antidiabetic Effects of Quercetin in Streptozocin-Induced Diabetic RatsDocument8 pagesAntidiabetic Effects of Quercetin in Streptozocin-Induced Diabetic RatsMohammad IvanNo ratings yet
- Vivo Analyses of Experimental Animals (6,7)Document23 pagesVivo Analyses of Experimental Animals (6,7)aakash sahaNo ratings yet
- 10.1515 - Labmed 2019 0098Document9 pages10.1515 - Labmed 2019 0098Fery ArdiansahNo ratings yet
- Cell Biochem Funct 2011 p55Document9 pagesCell Biochem Funct 2011 p55IQBAL HAKKIKINo ratings yet
- Extraction of Glycogen On Mild Condition Lacks Aig FractionDocument7 pagesExtraction of Glycogen On Mild Condition Lacks Aig FractionMaika DoronelaNo ratings yet
- Hyperglycemia As An Effect of Cardiopulmonary Bypass: Intra-Operative Glucose ManagementDocument6 pagesHyperglycemia As An Effect of Cardiopulmonary Bypass: Intra-Operative Glucose ManagementMuhammad RizqiNo ratings yet
- Liraglutide Improves Lipid Metabolism by Enhancing Cholesterol Efflux Associated With ABCA1 and ERK1/2 PathwayDocument12 pagesLiraglutide Improves Lipid Metabolism by Enhancing Cholesterol Efflux Associated With ABCA1 and ERK1/2 PathwayAndrianna NastasyaNo ratings yet
- Reti ManDocument8 pagesReti ManSUMIT PANDEYNo ratings yet
- Renal Threshold For Glucose ReabsorptionDocument4 pagesRenal Threshold For Glucose ReabsorptionsoniNo ratings yet
- Free Radical Biology and Medicine: Invited Review ArticleDocument13 pagesFree Radical Biology and Medicine: Invited Review ArticleNatalia SimõesNo ratings yet
- Re Glucose Reprt-1Document23 pagesRe Glucose Reprt-1Ingrid BayiyanaNo ratings yet
- Cancer Cell Get Fat From Glutamine 2012Document4 pagesCancer Cell Get Fat From Glutamine 2012Yulianti RiskiNo ratings yet
- ProImmune F1 On Adenine-Induced Chronic Progressive Tubulointerstitial NephropathyDocument12 pagesProImmune F1 On Adenine-Induced Chronic Progressive Tubulointerstitial NephropathyBryan Edson M. YcoNo ratings yet
- Methods: A Comparison of Three of Glycogen Measurement in TissuesDocument8 pagesMethods: A Comparison of Three of Glycogen Measurement in TissuesDawn KentNo ratings yet
- Ijms 21 07833Document25 pagesIjms 21 07833Zerrae CBTNo ratings yet
- Pradedova 2018Document9 pagesPradedova 2018rocio fernandez san juanNo ratings yet
- Glutathione Antioxidant Properties Dedicated To NaDocument21 pagesGlutathione Antioxidant Properties Dedicated To NaTazkiyatul Asma'iNo ratings yet
- A Novel Therapeutic Agent For Type 2 Diabetes Mellitus: SGLT2 InhibitorDocument13 pagesA Novel Therapeutic Agent For Type 2 Diabetes Mellitus: SGLT2 InhibitorSheldon SilvaNo ratings yet
- Sun, 2010 DHA Regulates Lipogenesis and Lipolysis Genes in Mice Adipose and LiverDocument7 pagesSun, 2010 DHA Regulates Lipogenesis and Lipolysis Genes in Mice Adipose and LiverDaniela Patricia Alvarez AravenaNo ratings yet
- HHS Public Access: Sodium-Glucose CotransportDocument12 pagesHHS Public Access: Sodium-Glucose CotransportAlexandra VásquezNo ratings yet
- MJMS30022023 09Document15 pagesMJMS30022023 09Muzammal RazaNo ratings yet
- CDD 2011102 ADocument12 pagesCDD 2011102 ACaerulus Fuad Abdul BaqiNo ratings yet
- 2.glycolysis & Oxidation of PyruvateDocument12 pages2.glycolysis & Oxidation of Pyruvateقتيبه خالد دحام خلفNo ratings yet
- Rompf Icfp15Document8 pagesRompf Icfp15MarrauNo ratings yet
- Tetrodotoxin: TTX: BackgroundDocument12 pagesTetrodotoxin: TTX: BackgroundMarrauNo ratings yet
- Royal FamilyDocument38 pagesRoyal FamilyMarrauNo ratings yet
- 2:4:6-Trinitrotoluene: 0-0114N-Ba (OH)Document16 pages2:4:6-Trinitrotoluene: 0-0114N-Ba (OH)MarrauNo ratings yet
- Penerapan Prinsip Rule of Reason Pada Putusan Perkara Nomor 08/KPPU-I/2020 Tentang Dugaan Praktik Diskriminasi Antara Telkom-Telkomsel Dan NetflixDocument14 pagesPenerapan Prinsip Rule of Reason Pada Putusan Perkara Nomor 08/KPPU-I/2020 Tentang Dugaan Praktik Diskriminasi Antara Telkom-Telkomsel Dan NetflixMarrauNo ratings yet
- Examining Explosives Handling Sensitivity of Trinitrotoluene (TNT) With Different Particle SizesDocument7 pagesExamining Explosives Handling Sensitivity of Trinitrotoluene (TNT) With Different Particle SizesMarrauNo ratings yet
- Toxins 14 00576 v2Document34 pagesToxins 14 00576 v2MarrauNo ratings yet
- Molecules TNTDocument9 pagesMolecules TNTMarrauNo ratings yet
- Q. Exp.: A To NerveDocument10 pagesQ. Exp.: A To NerveMarrauNo ratings yet
- Effect Salicylic Acid To LettuceDocument6 pagesEffect Salicylic Acid To LettuceMarrauNo ratings yet
- Aspirin: Damocles Gruppe C Daniel Deckenbach Bilal Danisman Benedict Depp Scotty CobbDocument7 pagesAspirin: Damocles Gruppe C Daniel Deckenbach Bilal Danisman Benedict Depp Scotty CobbMarrauNo ratings yet
- Partial Characterization of Tetrodotoxin-Binding Component From Nerve MembraneDocument4 pagesPartial Characterization of Tetrodotoxin-Binding Component From Nerve MembraneMarrauNo ratings yet
- ICRI Sunscreen 2Document21 pagesICRI Sunscreen 2MarrauNo ratings yet
- The Synthesis, Properties, & Application of Salicylic Acid in The Treatment of AcneDocument13 pagesThe Synthesis, Properties, & Application of Salicylic Acid in The Treatment of AcneMarrauNo ratings yet
- Biosyntesis of RetinolDocument5 pagesBiosyntesis of RetinolMarrauNo ratings yet
- Treatment of PainDocument5 pagesTreatment of PainMarrauNo ratings yet
- Breast CancerDocument36 pagesBreast CancerMarrauNo ratings yet
- Vitamin B3 at A Glance: CancerDocument3 pagesVitamin B3 at A Glance: CancerMarrauNo ratings yet
- Nutritional Medicine and Diet Care: ClinmedDocument5 pagesNutritional Medicine and Diet Care: ClinmedMarrauNo ratings yet
- Salicylic AcidDocument5 pagesSalicylic AcidMarrauNo ratings yet
- Retinol Product InformationDocument3 pagesRetinol Product InformationMarrauNo ratings yet
- It First Into Retinoic Acid. Retinyl Palmitate: Layers Up, It Is Safe To Use With AhasDocument2 pagesIt First Into Retinoic Acid. Retinyl Palmitate: Layers Up, It Is Safe To Use With AhasMarrauNo ratings yet
- 2dot3 AgingSkin PhysRes1 BellemereGDocument10 pages2dot3 AgingSkin PhysRes1 BellemereGMarrauNo ratings yet
- Outpatient Acne Care Guideline: SeverityDocument5 pagesOutpatient Acne Care Guideline: SeverityMarrauNo ratings yet
- Public Law 113-195 113th Congress An ActDocument18 pagesPublic Law 113-195 113th Congress An ActMarrauNo ratings yet
- Retinol in CosmeticsDocument204 pagesRetinol in CosmeticsMarrauNo ratings yet
- Sunscreen Market AnalysisDocument43 pagesSunscreen Market AnalysisMarrauNo ratings yet
- En515 2017Document11 pagesEn515 2017katemizNo ratings yet
- Aromatic Reactions ExercisesDocument6 pagesAromatic Reactions Exercisesisaacrenzie.dipus.sciNo ratings yet
- 100 MTPD Acetic Acid Plant DesignDocument60 pages100 MTPD Acetic Acid Plant Designmudasserulbuet18No ratings yet
- MCQs of Atomic and Nuclear Physics With AnswersDocument8 pagesMCQs of Atomic and Nuclear Physics With Answers09whitedevil90No ratings yet
- Tech Sheet - ProSilver45TDocument2 pagesTech Sheet - ProSilver45TMicheal Booka WhelanNo ratings yet
- 9701_m24_qp_42Document28 pages9701_m24_qp_42rashya.sharmaNo ratings yet
- RIL J4 Projects 2Document25 pagesRIL J4 Projects 2sumit6singhNo ratings yet
- General ToxicologyDocument28 pagesGeneral Toxicologyhhes8116No ratings yet
- Natural PolymersDocument35 pagesNatural Polymerssiti sunarintyasNo ratings yet
- Safety Data Sheet of XI40211-000Document11 pagesSafety Data Sheet of XI40211-000irvingrscNo ratings yet
- ALTA 2020 NCC - Queensland Pacific MetalsDocument19 pagesALTA 2020 NCC - Queensland Pacific MetalssirbrianmiloNo ratings yet
- Agilent 5100 Series ICP-OES Consumables Ordering GuideDocument44 pagesAgilent 5100 Series ICP-OES Consumables Ordering Guideandri purnomoNo ratings yet
- Method 6.0 2014Document2 pagesMethod 6.0 2014FelisbertoNo ratings yet
- Chem 1008 Lab Midterms 2Document6 pagesChem 1008 Lab Midterms 2Jie Ann Faith AusmoloNo ratings yet
- Early Stage Recovery of Lithium From Tailored Thermal Conditioned Black Mass Part I (Schwich Et Al 2021)Document30 pagesEarly Stage Recovery of Lithium From Tailored Thermal Conditioned Black Mass Part I (Schwich Et Al 2021)irdantoNo ratings yet
- Practice Test-02 Arjuna NEET (2025) 09.06.2024 SolutionsDocument15 pagesPractice Test-02 Arjuna NEET (2025) 09.06.2024 Solutionskenyen.jarekNo ratings yet
- Ehudin Jacs ironIVoxoDocument19 pagesEhudin Jacs ironIVoxoneerajNo ratings yet
- MSDS MotorexDocument6 pagesMSDS MotorexGezy MegalittaNo ratings yet
- COA Soda Ash Dense Ansac - 230522 - 143613Document2 pagesCOA Soda Ash Dense Ansac - 230522 - 143613nur.hudaNo ratings yet
- Honeywell Genetron R410aDocument15 pagesHoneywell Genetron R410avivekNo ratings yet
- Yosim and MayerDocument66 pagesYosim and MayerRajabMumbeeNo ratings yet
- Ebook Chemistry For Today General Organic and Biochemistry PDF Full Chapter PDFDocument67 pagesEbook Chemistry For Today General Organic and Biochemistry PDF Full Chapter PDFrobert.davidson233100% (31)
- Ams 4911LDocument7 pagesAms 4911Ljamesdong1120No ratings yet
- Calculation Part 2 in AC-2Document2 pagesCalculation Part 2 in AC-223005852No ratings yet
- Grade 12 P2 2016 - 2023 papersDocument287 pagesGrade 12 P2 2016 - 2023 paperslihlemananga65No ratings yet
- Ebook Corrosion Atlas A Collection of Illustrated Case Histories PDF Full Chapter PDFDocument55 pagesEbook Corrosion Atlas A Collection of Illustrated Case Histories PDF Full Chapter PDFedwina.perry203100% (40)
- Module 1 (Thermo 2)Document38 pagesModule 1 (Thermo 2)Genesis MedelNo ratings yet
- KCET 2024 Chemistry Paper With AnswerDocument9 pagesKCET 2024 Chemistry Paper With Answerthejasmath2005No ratings yet
- CZ 132Document1 pageCZ 132Taner DemirelNo ratings yet
- Aldehydes, Ketones, and Carboxylic Acids Shobhit NirwanDocument12 pagesAldehydes, Ketones, and Carboxylic Acids Shobhit Nirwangokulss374100% (1)