
Louca 2016 Global Ocean
Louca 2016 Global Ocean
You might also like
- Great Apes Anticipate That Other Individuals Will Act According To False BeliefsDocument6 pagesGreat Apes Anticipate That Other Individuals Will Act According To False BeliefsjavieraNo ratings yet
- Sarafian 2014Document5 pagesSarafian 2014mariano totoNo ratings yet
- 4D Electron Tomography ScienceDocument6 pages4D Electron Tomography Sciencehappynewyear11No ratings yet
- Chapter - 4 Carbon NanotubesDocument8 pagesChapter - 4 Carbon NanotubeskarthikNo ratings yet
- Banerjee 2008Document6 pagesBanerjee 2008paulaNo ratings yet
- Visualizing Individual Nitrogen Dopants in Monolayer GrapheneDocument6 pagesVisualizing Individual Nitrogen Dopants in Monolayer Graphene김영찬No ratings yet
- 623 FullDocument5 pages623 Fullmariano totoNo ratings yet
- A Reconciled Estimate of Ice-Sheet Mass Balance Shepherd Et Al 2012Document8 pagesA Reconciled Estimate of Ice-Sheet Mass Balance Shepherd Et Al 2012silvabussoNo ratings yet
- Chalfie 1994Document4 pagesChalfie 1994Basab BijayeeNo ratings yet
- Spinal Cord Stimulation Restores Locomotion in Animal Models of Parkinson S DiseaseDocument6 pagesSpinal Cord Stimulation Restores Locomotion in Animal Models of Parkinson S DiseaseDiego Eduardo HurtadoNo ratings yet
- Science 1257158Document4 pagesScience 12571581592162022No ratings yet
- Spatiotemporal Microbial Evolution On Antibiotic Landscapes: 13 ReferencesandnotesDocument6 pagesSpatiotemporal Microbial Evolution On Antibiotic Landscapes: 13 ReferencesandnotesJaqueline Godinez CamachoNo ratings yet
- Spatiotemporal Microbial Evolution On Antibiotic Landscapes: 13 ReferencesandnotesDocument6 pagesSpatiotemporal Microbial Evolution On Antibiotic Landscapes: 13 ReferencesandnotesFranco VeglianiNo ratings yet
- Expanding The Fluorine Chemistry of Living Systems Using Engineered Polyketide Synthase PathwaysDocument7 pagesExpanding The Fluorine Chemistry of Living Systems Using Engineered Polyketide Synthase PathwaysJunia Alfa NessaNo ratings yet
- Nanoscale Tunable Reduction of Graphene Oxide For Graphene ElectronicsDocument6 pagesNanoscale Tunable Reduction of Graphene Oxide For Graphene Electronicsadrianoreis1No ratings yet
- Barriopedro 11Document6 pagesBarriopedro 11JxPNo ratings yet
- Oso PolarDocument5 pagesOso PolarGiovanni Ávila EsquivelNo ratings yet
- A Four-Legged Snake From The Early Cretaceous of GondwanaDocument6 pagesA Four-Legged Snake From The Early Cretaceous of GondwanaYilbert Oswaldo Jimenez CanoNo ratings yet
- 2002 Irmof-2 EdusolDocument5 pages2002 Irmof-2 EdusolAlejandra AwimbaweNo ratings yet
- 2002 Irmof-1 EdusifDocument5 pages2002 Irmof-1 EdusifAlejandra AwimbaweNo ratings yet
- Just Think: The Challenges of The Disengaged MindDocument4 pagesJust Think: The Challenges of The Disengaged MindRaja SelvarajanNo ratings yet
- Wu Canopy Phenology Photosythesis Seasonality Amazonia 2016Document6 pagesWu Canopy Phenology Photosythesis Seasonality Amazonia 2016Lucas BauerNo ratings yet
- BonachelaDocument6 pagesBonachelaSuane SouzaNo ratings yet
- Science 1202393Document8 pagesScience 1202393CARLOS FRANCISCO MANTILLA MONTALVONo ratings yet
- Ref 1Document5 pagesRef 1Diego Alejandro Roa CalaNo ratings yet
- 2007 Ultrastrong and Stiff Layered Polymer Nanocomposites ScienceDocument5 pages2007 Ultrastrong and Stiff Layered Polymer Nanocomposites ScienceНаталья ГайдукNo ratings yet
- He Cken Berger 92008Document6 pagesHe Cken Berger 92008llerenasNo ratings yet
- Exclusion HurtDocument4 pagesExclusion HurtMarina Sánchez PicazoNo ratings yet
- Self-Assembly and Mineralization of Peptide-Amphiphile NanofibersDocument5 pagesSelf-Assembly and Mineralization of Peptide-Amphiphile NanofibersJosé Rodrigo Alejandro Martínez DíazNo ratings yet
- DiversityDocument4 pagesDiversityFernando BiscioneNo ratings yet
- Thermally Stable Single-Atom Platinum-On-Ceria Catalysts Via Atom TrappingDocument6 pagesThermally Stable Single-Atom Platinum-On-Ceria Catalysts Via Atom Trappingsergioodin4851No ratings yet
- Daley Et Al 2009Document4 pagesDaley Et Al 2009SedPaleoNo ratings yet
- Genomic RemodellingDocument2 pagesGenomic RemodellingDenys Quispe AlcocerNo ratings yet
- Improved Oxygen Reduction Activity On Pt3Ni Via Increased Surface Site AvailabilityDocument6 pagesImproved Oxygen Reduction Activity On Pt3Ni Via Increased Surface Site AvailabilityARIADNANo ratings yet
- Mesotunnels On The Silica Wall of Ordered SBA-15 To Generate Three-Dimensional Large-Pore Mesoporous NetworksDocument2 pagesMesotunnels On The Silica Wall of Ordered SBA-15 To Generate Three-Dimensional Large-Pore Mesoporous NetworksAneesh KumarNo ratings yet
- Health Benefits of Eating Chocolate?: Brief CommunicationsDocument2 pagesHealth Benefits of Eating Chocolate?: Brief CommunicationsAimée SanchezNo ratings yet
- 5-2013-Research-Multiplex Genome Engineering Using CRISPRCas SystemsDocument6 pages5-2013-Research-Multiplex Genome Engineering Using CRISPRCas SystemsCristian Felipe Sandoval QuiñonezNo ratings yet
- A General Mechanism For Network-Dosage Compensation in Gene CircuitsDocument6 pagesA General Mechanism For Network-Dosage Compensation in Gene CircuitsVijendra KavatalkarNo ratings yet
- A Porous Silicon-Based Optical Interferometric BiosensorDocument5 pagesA Porous Silicon-Based Optical Interferometric BiosensorTahmid Hassan TalukdarNo ratings yet
- Porous Silicon BiosensorDocument5 pagesPorous Silicon BiosensorTahmid Hassan TalukdarNo ratings yet
- Science 1213229Document5 pagesScience 12132291592162022No ratings yet
- Multiwavelength Achromatic Metasurfaces by Dispersive Phase CompensationDocument5 pagesMultiwavelength Achromatic Metasurfaces by Dispersive Phase CompensationTahmid Hassan TalukdarNo ratings yet
- Quantom Dot Induced Phase Stabilization of The Cspbi3 Perovskite For High Efficiency PhotovoltaicsDocument5 pagesQuantom Dot Induced Phase Stabilization of The Cspbi3 Perovskite For High Efficiency PhotovoltaicsabuhurairabscNo ratings yet
- Angew Chem Int Ed - 2000 - Taguchi - Living Coordination Polymerization of Allene Derivatives Bearing Hydroxy Groups byDocument3 pagesAngew Chem Int Ed - 2000 - Taguchi - Living Coordination Polymerization of Allene Derivatives Bearing Hydroxy Groups byXIAJIE WANGNo ratings yet
- Wu Et Al. - 2013 - Science (New York, N.Y.)Document6 pagesWu Et Al. - 2013 - Science (New York, N.Y.)Mariana NannettiNo ratings yet
- 2005 NatureDocument5 pages2005 NaturecrackintheshatNo ratings yet
- Crystal Plane Dependent Growth of Aligned Single-Walled Carbon Nanotubes On SapphireDocument8 pagesCrystal Plane Dependent Growth of Aligned Single-Walled Carbon Nanotubes On SapphireDiavolo RequiemNo ratings yet
- Just Think: The Challenges of The Disengaged MindDocument29 pagesJust Think: The Challenges of The Disengaged MindArjun BalajiNo ratings yet
- Dhuime Et Al. 2012 - ScienceDocument4 pagesDhuime Et Al. 2012 - ScienceLady Johana Buritica CortesNo ratings yet
- Mollusca Li Et Al 2015 Biomineralized Armor Chiton EyeDocument6 pagesMollusca Li Et Al 2015 Biomineralized Armor Chiton EyeAlejandro MontalvoNo ratings yet
- 2D LnIIIRuIII2 Compounds Constructed From Trans - (Ru (Acac) 2 (CN) 2) - . Syntheses, Structures, and Magnetic PropertiesDocument5 pages2D LnIIIRuIII2 Compounds Constructed From Trans - (Ru (Acac) 2 (CN) 2) - . Syntheses, Structures, and Magnetic PropertiesKamal Uddin AnsariNo ratings yet
- Photonic Band Gap Guidance in Optical Fibers: J. C. Knight, J. Broeng, T. A. Birks, P. St. J. RussellDocument3 pagesPhotonic Band Gap Guidance in Optical Fibers: J. C. Knight, J. Broeng, T. A. Birks, P. St. J. RussellArnab BhowmikNo ratings yet
- Argonauta 3Document6 pagesArgonauta 3Christian Moises CasasNo ratings yet
- 2010 Domain Dynamics During Ferroelectric SwitchingDocument6 pages2010 Domain Dynamics During Ferroelectric SwitchingAnaghaNo ratings yet
- Proyecto FisicoquimicaDocument5 pagesProyecto FisicoquimicaRosa LozanoNo ratings yet
- Small-Diameter Silicon Nanowire Surfaces: EportsDocument5 pagesSmall-Diameter Silicon Nanowire Surfaces: EportsBeatrice ChouNo ratings yet
- X-Ray Scattering Study of Thin Films of Poly (2,5-Bis (3-Alkylthiophen-2-Yl) Thieno (3,2-) Thiophene)Document12 pagesX-Ray Scattering Study of Thin Films of Poly (2,5-Bis (3-Alkylthiophen-2-Yl) Thieno (3,2-) Thiophene)kastorNo ratings yet
- Science 282 5386 98Document3 pagesScience 282 5386 98scribduserme123No ratings yet
- The Smallest Carbon Nanotube: Brief CommunicationsDocument2 pagesThe Smallest Carbon Nanotube: Brief CommunicationsSon NguyenNo ratings yet
- Anomalous Rare Earths and Actinides: Valence Fluctuation and Heavy FermionsFrom EverandAnomalous Rare Earths and Actinides: Valence Fluctuation and Heavy FermionsJ.X. BoucherleNo ratings yet
- Otolith Function and DisordersDocument139 pagesOtolith Function and DisordersAlbert Gheorghe100% (1)
- Silk'n Lipo - Clinical OverviewDocument10 pagesSilk'n Lipo - Clinical OverviewFit BoxxNo ratings yet
- How Cancer Spreads MetastasisDocument6 pagesHow Cancer Spreads MetastasisAfia TawiahNo ratings yet
- Basic Concepts in PharmacologyDocument25 pagesBasic Concepts in PharmacologyRizka Amaliah NasutionNo ratings yet
- Bcsci 10 Unit 1 Topic1 2Document20 pagesBcsci 10 Unit 1 Topic1 2DemisNo ratings yet
- Genetic Disorders Project Rubric 17-18Document3 pagesGenetic Disorders Project Rubric 17-18api-394564312No ratings yet
- Chap 19 EditDocument16 pagesChap 19 EditSherlock Wesley ConanNo ratings yet
- A New 5,6-Dihydro-2-Pyrone Derivative From Phomopsis Amygdali, An Endophytic Fungus Isolated From Hazelnut (Corylus Avellana)Document4 pagesA New 5,6-Dihydro-2-Pyrone Derivative From Phomopsis Amygdali, An Endophytic Fungus Isolated From Hazelnut (Corylus Avellana)akayserefNo ratings yet
- 73-Article Text-213-1-10-20180820Document5 pages73-Article Text-213-1-10-20180820Putri HaniahNo ratings yet
- ATC Lab Set UpDocument84 pagesATC Lab Set UpKrithika BalasubramanianNo ratings yet
- HemostasisDocument5 pagesHemostasisPadmavathi C100% (1)
- Provisional: October 2021 Examination TimetableDocument13 pagesProvisional: October 2021 Examination TimetableNicholson Nicholson0% (1)
- Unlocking The Potential of Orphan Legumes: Christopher Cullis and Karl J. KunertDocument9 pagesUnlocking The Potential of Orphan Legumes: Christopher Cullis and Karl J. KunertKaniNo ratings yet
- 1st Periodical TestDocument8 pages1st Periodical TestJunard AsentistaNo ratings yet
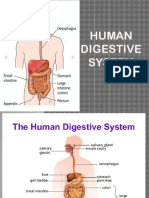
- Digestive-System-2 Grade 8Document39 pagesDigestive-System-2 Grade 8Joshua Muego100% (2)
- (Current Topics in Membranes 72) Vann Bennett (Eds.) - Functional Organization of Vertebrate Plasma Membrane-Academic Press (2013) PDFDocument364 pages(Current Topics in Membranes 72) Vann Bennett (Eds.) - Functional Organization of Vertebrate Plasma Membrane-Academic Press (2013) PDFSankarshan talluriNo ratings yet
- June 2017 (v2) MS - Paper 4 CIE Biology IGCSEDocument11 pagesJune 2017 (v2) MS - Paper 4 CIE Biology IGCSERahulBansumanNo ratings yet
- Lesson Title: Digestion/Absorption of Nutrients: 7 Grade General Science CourseDocument5 pagesLesson Title: Digestion/Absorption of Nutrients: 7 Grade General Science CourserebbiegNo ratings yet
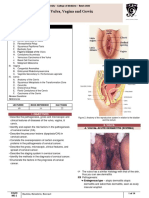
- B14PathologyL1 - Diseases of The Vulva, Vaginal, and CervixDocument14 pagesB14PathologyL1 - Diseases of The Vulva, Vaginal, and CervixBarda GulanNo ratings yet
- Name: - Section: - Schedule: - Class Number: - DateDocument8 pagesName: - Section: - Schedule: - Class Number: - DateDalrymple Clavo RamosNo ratings yet
- Paper - Designer BabyDocument13 pagesPaper - Designer Babyary sofyanNo ratings yet
- Leukocytes 2Document19 pagesLeukocytes 2Tabitha EaglehouseNo ratings yet
- Classification of EnzymesDocument7 pagesClassification of EnzymesAbhik ShrivastavaNo ratings yet
- Duchenne Muscular Dystrophy: June 2014Document6 pagesDuchenne Muscular Dystrophy: June 2014Beta Yunita PNo ratings yet
- Micro LectureDocument4 pagesMicro LectureJewel OlaezNo ratings yet
- تجميعات باثولوجيDocument3 pagesتجميعات باثولوجيTurky TurkyNo ratings yet
- Microbiologist or Lab TechnicianDocument2 pagesMicrobiologist or Lab Technicianapi-78130315No ratings yet
- The Chicken Genetic Family Tree For PrintingDocument2 pagesThe Chicken Genetic Family Tree For PrintingDebbie Samar BoragayNo ratings yet
- Introduction To ParasitologyDocument36 pagesIntroduction To ParasitologyMj BrionesNo ratings yet
- BIO (Study Guide)Document15 pagesBIO (Study Guide)Izzy Dynielle SolamilloNo ratings yet
























































































Download as pdf or txt
You might also like
- Great Apes Anticipate That Other Individuals Will Act According To False BeliefsDocument6 pagesGreat Apes Anticipate That Other Individuals Will Act According To False BeliefsjavieraNo ratings yet
- Sarafian 2014Document5 pagesSarafian 2014mariano totoNo ratings yet
- 4D Electron Tomography ScienceDocument6 pages4D Electron Tomography Sciencehappynewyear11No ratings yet
- Chapter - 4 Carbon NanotubesDocument8 pagesChapter - 4 Carbon NanotubeskarthikNo ratings yet
- Banerjee 2008Document6 pagesBanerjee 2008paulaNo ratings yet
- Visualizing Individual Nitrogen Dopants in Monolayer GrapheneDocument6 pagesVisualizing Individual Nitrogen Dopants in Monolayer Graphene김영찬No ratings yet
- 623 FullDocument5 pages623 Fullmariano totoNo ratings yet
- A Reconciled Estimate of Ice-Sheet Mass Balance Shepherd Et Al 2012Document8 pagesA Reconciled Estimate of Ice-Sheet Mass Balance Shepherd Et Al 2012silvabussoNo ratings yet
- Chalfie 1994Document4 pagesChalfie 1994Basab BijayeeNo ratings yet
- Spinal Cord Stimulation Restores Locomotion in Animal Models of Parkinson S DiseaseDocument6 pagesSpinal Cord Stimulation Restores Locomotion in Animal Models of Parkinson S DiseaseDiego Eduardo HurtadoNo ratings yet
- Science 1257158Document4 pagesScience 12571581592162022No ratings yet
- Spatiotemporal Microbial Evolution On Antibiotic Landscapes: 13 ReferencesandnotesDocument6 pagesSpatiotemporal Microbial Evolution On Antibiotic Landscapes: 13 ReferencesandnotesJaqueline Godinez CamachoNo ratings yet
- Spatiotemporal Microbial Evolution On Antibiotic Landscapes: 13 ReferencesandnotesDocument6 pagesSpatiotemporal Microbial Evolution On Antibiotic Landscapes: 13 ReferencesandnotesFranco VeglianiNo ratings yet
- Expanding The Fluorine Chemistry of Living Systems Using Engineered Polyketide Synthase PathwaysDocument7 pagesExpanding The Fluorine Chemistry of Living Systems Using Engineered Polyketide Synthase PathwaysJunia Alfa NessaNo ratings yet
- Nanoscale Tunable Reduction of Graphene Oxide For Graphene ElectronicsDocument6 pagesNanoscale Tunable Reduction of Graphene Oxide For Graphene Electronicsadrianoreis1No ratings yet
- Barriopedro 11Document6 pagesBarriopedro 11JxPNo ratings yet
- Oso PolarDocument5 pagesOso PolarGiovanni Ávila EsquivelNo ratings yet
- A Four-Legged Snake From The Early Cretaceous of GondwanaDocument6 pagesA Four-Legged Snake From The Early Cretaceous of GondwanaYilbert Oswaldo Jimenez CanoNo ratings yet
- 2002 Irmof-2 EdusolDocument5 pages2002 Irmof-2 EdusolAlejandra AwimbaweNo ratings yet
- 2002 Irmof-1 EdusifDocument5 pages2002 Irmof-1 EdusifAlejandra AwimbaweNo ratings yet
- Just Think: The Challenges of The Disengaged MindDocument4 pagesJust Think: The Challenges of The Disengaged MindRaja SelvarajanNo ratings yet
- Wu Canopy Phenology Photosythesis Seasonality Amazonia 2016Document6 pagesWu Canopy Phenology Photosythesis Seasonality Amazonia 2016Lucas BauerNo ratings yet
- BonachelaDocument6 pagesBonachelaSuane SouzaNo ratings yet
- Science 1202393Document8 pagesScience 1202393CARLOS FRANCISCO MANTILLA MONTALVONo ratings yet
- Ref 1Document5 pagesRef 1Diego Alejandro Roa CalaNo ratings yet
- 2007 Ultrastrong and Stiff Layered Polymer Nanocomposites ScienceDocument5 pages2007 Ultrastrong and Stiff Layered Polymer Nanocomposites ScienceНаталья ГайдукNo ratings yet
- He Cken Berger 92008Document6 pagesHe Cken Berger 92008llerenasNo ratings yet
- Exclusion HurtDocument4 pagesExclusion HurtMarina Sánchez PicazoNo ratings yet
- Self-Assembly and Mineralization of Peptide-Amphiphile NanofibersDocument5 pagesSelf-Assembly and Mineralization of Peptide-Amphiphile NanofibersJosé Rodrigo Alejandro Martínez DíazNo ratings yet
- DiversityDocument4 pagesDiversityFernando BiscioneNo ratings yet
- Thermally Stable Single-Atom Platinum-On-Ceria Catalysts Via Atom TrappingDocument6 pagesThermally Stable Single-Atom Platinum-On-Ceria Catalysts Via Atom Trappingsergioodin4851No ratings yet
- Daley Et Al 2009Document4 pagesDaley Et Al 2009SedPaleoNo ratings yet
- Genomic RemodellingDocument2 pagesGenomic RemodellingDenys Quispe AlcocerNo ratings yet
- Improved Oxygen Reduction Activity On Pt3Ni Via Increased Surface Site AvailabilityDocument6 pagesImproved Oxygen Reduction Activity On Pt3Ni Via Increased Surface Site AvailabilityARIADNANo ratings yet
- Mesotunnels On The Silica Wall of Ordered SBA-15 To Generate Three-Dimensional Large-Pore Mesoporous NetworksDocument2 pagesMesotunnels On The Silica Wall of Ordered SBA-15 To Generate Three-Dimensional Large-Pore Mesoporous NetworksAneesh KumarNo ratings yet
- Health Benefits of Eating Chocolate?: Brief CommunicationsDocument2 pagesHealth Benefits of Eating Chocolate?: Brief CommunicationsAimée SanchezNo ratings yet
- 5-2013-Research-Multiplex Genome Engineering Using CRISPRCas SystemsDocument6 pages5-2013-Research-Multiplex Genome Engineering Using CRISPRCas SystemsCristian Felipe Sandoval QuiñonezNo ratings yet
- A General Mechanism For Network-Dosage Compensation in Gene CircuitsDocument6 pagesA General Mechanism For Network-Dosage Compensation in Gene CircuitsVijendra KavatalkarNo ratings yet
- A Porous Silicon-Based Optical Interferometric BiosensorDocument5 pagesA Porous Silicon-Based Optical Interferometric BiosensorTahmid Hassan TalukdarNo ratings yet
- Porous Silicon BiosensorDocument5 pagesPorous Silicon BiosensorTahmid Hassan TalukdarNo ratings yet
- Science 1213229Document5 pagesScience 12132291592162022No ratings yet
- Multiwavelength Achromatic Metasurfaces by Dispersive Phase CompensationDocument5 pagesMultiwavelength Achromatic Metasurfaces by Dispersive Phase CompensationTahmid Hassan TalukdarNo ratings yet
- Quantom Dot Induced Phase Stabilization of The Cspbi3 Perovskite For High Efficiency PhotovoltaicsDocument5 pagesQuantom Dot Induced Phase Stabilization of The Cspbi3 Perovskite For High Efficiency PhotovoltaicsabuhurairabscNo ratings yet
- Angew Chem Int Ed - 2000 - Taguchi - Living Coordination Polymerization of Allene Derivatives Bearing Hydroxy Groups byDocument3 pagesAngew Chem Int Ed - 2000 - Taguchi - Living Coordination Polymerization of Allene Derivatives Bearing Hydroxy Groups byXIAJIE WANGNo ratings yet
- Wu Et Al. - 2013 - Science (New York, N.Y.)Document6 pagesWu Et Al. - 2013 - Science (New York, N.Y.)Mariana NannettiNo ratings yet
- 2005 NatureDocument5 pages2005 NaturecrackintheshatNo ratings yet
- Crystal Plane Dependent Growth of Aligned Single-Walled Carbon Nanotubes On SapphireDocument8 pagesCrystal Plane Dependent Growth of Aligned Single-Walled Carbon Nanotubes On SapphireDiavolo RequiemNo ratings yet
- Just Think: The Challenges of The Disengaged MindDocument29 pagesJust Think: The Challenges of The Disengaged MindArjun BalajiNo ratings yet
- Dhuime Et Al. 2012 - ScienceDocument4 pagesDhuime Et Al. 2012 - ScienceLady Johana Buritica CortesNo ratings yet
- Mollusca Li Et Al 2015 Biomineralized Armor Chiton EyeDocument6 pagesMollusca Li Et Al 2015 Biomineralized Armor Chiton EyeAlejandro MontalvoNo ratings yet
- 2D LnIIIRuIII2 Compounds Constructed From Trans - (Ru (Acac) 2 (CN) 2) - . Syntheses, Structures, and Magnetic PropertiesDocument5 pages2D LnIIIRuIII2 Compounds Constructed From Trans - (Ru (Acac) 2 (CN) 2) - . Syntheses, Structures, and Magnetic PropertiesKamal Uddin AnsariNo ratings yet
- Photonic Band Gap Guidance in Optical Fibers: J. C. Knight, J. Broeng, T. A. Birks, P. St. J. RussellDocument3 pagesPhotonic Band Gap Guidance in Optical Fibers: J. C. Knight, J. Broeng, T. A. Birks, P. St. J. RussellArnab BhowmikNo ratings yet
- Argonauta 3Document6 pagesArgonauta 3Christian Moises CasasNo ratings yet
- 2010 Domain Dynamics During Ferroelectric SwitchingDocument6 pages2010 Domain Dynamics During Ferroelectric SwitchingAnaghaNo ratings yet
- Proyecto FisicoquimicaDocument5 pagesProyecto FisicoquimicaRosa LozanoNo ratings yet
- Small-Diameter Silicon Nanowire Surfaces: EportsDocument5 pagesSmall-Diameter Silicon Nanowire Surfaces: EportsBeatrice ChouNo ratings yet
- X-Ray Scattering Study of Thin Films of Poly (2,5-Bis (3-Alkylthiophen-2-Yl) Thieno (3,2-) Thiophene)Document12 pagesX-Ray Scattering Study of Thin Films of Poly (2,5-Bis (3-Alkylthiophen-2-Yl) Thieno (3,2-) Thiophene)kastorNo ratings yet
- Science 282 5386 98Document3 pagesScience 282 5386 98scribduserme123No ratings yet
- The Smallest Carbon Nanotube: Brief CommunicationsDocument2 pagesThe Smallest Carbon Nanotube: Brief CommunicationsSon NguyenNo ratings yet
- Anomalous Rare Earths and Actinides: Valence Fluctuation and Heavy FermionsFrom EverandAnomalous Rare Earths and Actinides: Valence Fluctuation and Heavy FermionsJ.X. BoucherleNo ratings yet
- Otolith Function and DisordersDocument139 pagesOtolith Function and DisordersAlbert Gheorghe100% (1)
- Silk'n Lipo - Clinical OverviewDocument10 pagesSilk'n Lipo - Clinical OverviewFit BoxxNo ratings yet
- How Cancer Spreads MetastasisDocument6 pagesHow Cancer Spreads MetastasisAfia TawiahNo ratings yet
- Basic Concepts in PharmacologyDocument25 pagesBasic Concepts in PharmacologyRizka Amaliah NasutionNo ratings yet
- Bcsci 10 Unit 1 Topic1 2Document20 pagesBcsci 10 Unit 1 Topic1 2DemisNo ratings yet
- Genetic Disorders Project Rubric 17-18Document3 pagesGenetic Disorders Project Rubric 17-18api-394564312No ratings yet
- Chap 19 EditDocument16 pagesChap 19 EditSherlock Wesley ConanNo ratings yet
- A New 5,6-Dihydro-2-Pyrone Derivative From Phomopsis Amygdali, An Endophytic Fungus Isolated From Hazelnut (Corylus Avellana)Document4 pagesA New 5,6-Dihydro-2-Pyrone Derivative From Phomopsis Amygdali, An Endophytic Fungus Isolated From Hazelnut (Corylus Avellana)akayserefNo ratings yet
- 73-Article Text-213-1-10-20180820Document5 pages73-Article Text-213-1-10-20180820Putri HaniahNo ratings yet
- ATC Lab Set UpDocument84 pagesATC Lab Set UpKrithika BalasubramanianNo ratings yet
- HemostasisDocument5 pagesHemostasisPadmavathi C100% (1)
- Provisional: October 2021 Examination TimetableDocument13 pagesProvisional: October 2021 Examination TimetableNicholson Nicholson0% (1)
- Unlocking The Potential of Orphan Legumes: Christopher Cullis and Karl J. KunertDocument9 pagesUnlocking The Potential of Orphan Legumes: Christopher Cullis and Karl J. KunertKaniNo ratings yet
- 1st Periodical TestDocument8 pages1st Periodical TestJunard AsentistaNo ratings yet
- Digestive-System-2 Grade 8Document39 pagesDigestive-System-2 Grade 8Joshua Muego100% (2)
- (Current Topics in Membranes 72) Vann Bennett (Eds.) - Functional Organization of Vertebrate Plasma Membrane-Academic Press (2013) PDFDocument364 pages(Current Topics in Membranes 72) Vann Bennett (Eds.) - Functional Organization of Vertebrate Plasma Membrane-Academic Press (2013) PDFSankarshan talluriNo ratings yet
- June 2017 (v2) MS - Paper 4 CIE Biology IGCSEDocument11 pagesJune 2017 (v2) MS - Paper 4 CIE Biology IGCSERahulBansumanNo ratings yet
- Lesson Title: Digestion/Absorption of Nutrients: 7 Grade General Science CourseDocument5 pagesLesson Title: Digestion/Absorption of Nutrients: 7 Grade General Science CourserebbiegNo ratings yet
- B14PathologyL1 - Diseases of The Vulva, Vaginal, and CervixDocument14 pagesB14PathologyL1 - Diseases of The Vulva, Vaginal, and CervixBarda GulanNo ratings yet
- Name: - Section: - Schedule: - Class Number: - DateDocument8 pagesName: - Section: - Schedule: - Class Number: - DateDalrymple Clavo RamosNo ratings yet
- Paper - Designer BabyDocument13 pagesPaper - Designer Babyary sofyanNo ratings yet
- Leukocytes 2Document19 pagesLeukocytes 2Tabitha EaglehouseNo ratings yet
- Classification of EnzymesDocument7 pagesClassification of EnzymesAbhik ShrivastavaNo ratings yet
- Duchenne Muscular Dystrophy: June 2014Document6 pagesDuchenne Muscular Dystrophy: June 2014Beta Yunita PNo ratings yet
- Micro LectureDocument4 pagesMicro LectureJewel OlaezNo ratings yet
- تجميعات باثولوجيDocument3 pagesتجميعات باثولوجيTurky TurkyNo ratings yet
- Microbiologist or Lab TechnicianDocument2 pagesMicrobiologist or Lab Technicianapi-78130315No ratings yet
- The Chicken Genetic Family Tree For PrintingDocument2 pagesThe Chicken Genetic Family Tree For PrintingDebbie Samar BoragayNo ratings yet
- Introduction To ParasitologyDocument36 pagesIntroduction To ParasitologyMj BrionesNo ratings yet
- BIO (Study Guide)Document15 pagesBIO (Study Guide)Izzy Dynielle SolamilloNo ratings yet