Download as pdf or txt
You might also like
- Bioinformatics and Functional Genomics Ebook PDF VersionDocument61 pagesBioinformatics and Functional Genomics Ebook PDF Versionramona.evans546100% (53)
- F G Priest & I Cambell - Brewing Microbiology PDFDocument312 pagesF G Priest & I Cambell - Brewing Microbiology PDFOrlando Gonzalez100% (1)
- Biofuels: Alternative Feedstocks and Conversion ProcessesFrom EverandBiofuels: Alternative Feedstocks and Conversion ProcessesRating: 3 out of 5 stars3/5 (1)
- The Dog GenomeDocument21 pagesThe Dog Genomediparayogagalih100% (1)
- C Metabolic Flux Analysis-Guided Metabolic Engineering of Escherichia Coli For Improved Acetol Production From GlycerolDocument13 pagesC Metabolic Flux Analysis-Guided Metabolic Engineering of Escherichia Coli For Improved Acetol Production From GlycerolHAMMERLY JONATHAN LINO FUENTES RIVERANo ratings yet
- Yarrowia Lipolytica - Art. ScienceDirect - Kuttuvan Valappil Sanja,... Ashok Pandey (2015)Document19 pagesYarrowia Lipolytica - Art. ScienceDirect - Kuttuvan Valappil Sanja,... Ashok Pandey (2015)Regi RGNo ratings yet
- Bioproduction of Butanol From Biomass: From Genes To BioreactorsDocument8 pagesBioproduction of Butanol From Biomass: From Genes To BioreactorsOana VasileNo ratings yet
- Bioengineering: Converting Sugars To Biofuels: Ethanol and BeyondDocument20 pagesBioengineering: Converting Sugars To Biofuels: Ethanol and BeyondprosperNo ratings yet
- UAS - Paper Simulasi - Kelompok 6Document14 pagesUAS - Paper Simulasi - Kelompok 6Aryo RagaNo ratings yet
- (Atsumi, S. Hanai, T. Liao, James C., 2008) Non-Fermentative Pathways For Synthesis of Brainched-Chain Higher Alcohols As Biofuels'. Nature. No. 451, Pp. 86-90Document5 pages(Atsumi, S. Hanai, T. Liao, James C., 2008) Non-Fermentative Pathways For Synthesis of Brainched-Chain Higher Alcohols As Biofuels'. Nature. No. 451, Pp. 86-90Anastasia Ayu PratiwiNo ratings yet
- Alkaline Organosolv Pretreatment of Different Sorghum Stem Parts For Enhancing The Total Reducing Sugar Yields and P-Coumaric Acid ReleaseDocument14 pagesAlkaline Organosolv Pretreatment of Different Sorghum Stem Parts For Enhancing The Total Reducing Sugar Yields and P-Coumaric Acid Releaseuliseshbq7No ratings yet
- Dandan Li, Liangkun Long Dan Shaojun Ding. 2020. Alkaline Organosolv Pretreatment of DifferentDocument14 pagesDandan Li, Liangkun Long Dan Shaojun Ding. 2020. Alkaline Organosolv Pretreatment of DifferentAdhan AkbarNo ratings yet
- Manipulation of Malic Enzyme in Saccharomyces Cerevisiae For Increasing NADPH ProductionDocument12 pagesManipulation of Malic Enzyme in Saccharomyces Cerevisiae For Increasing NADPH ProductionJesús Miguel Jacinto NavaNo ratings yet
- A Simple Metabolic Ux Balance Analysis of Biomass and Bioethanol Production in Saccharomyces Cerevisiae Fed-Batch CulturesDocument11 pagesA Simple Metabolic Ux Balance Analysis of Biomass and Bioethanol Production in Saccharomyces Cerevisiae Fed-Batch Culturesmuhammad azizul hakimNo ratings yet
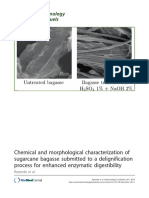
- Chemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityDocument19 pagesChemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityTabiita Huaman NorabuenaNo ratings yet
- BMC Biotechnology: Biosynthesis of Plant-Specific Stilbene Polyketides in Metabolically Engineered Escherichia ColiDocument12 pagesBMC Biotechnology: Biosynthesis of Plant-Specific Stilbene Polyketides in Metabolically Engineered Escherichia Colifather45No ratings yet
- Continuous and Batch Cultures of Escherichia Coli KJ134 For Succinic Acid Fermentation: Metabolic Flux Distributions and Production CharacteristicsDocument10 pagesContinuous and Batch Cultures of Escherichia Coli KJ134 For Succinic Acid Fermentation: Metabolic Flux Distributions and Production CharacteristicsSyed ShahNo ratings yet
- Cheng 2017Document14 pagesCheng 2017Insanul KholikNo ratings yet
- Lactic Acid Production Using Cheese Whey Based Medium in A Stirred Tank Reactor by A Ccpa Mutant of Lacticaseibacillus CaseiDocument13 pagesLactic Acid Production Using Cheese Whey Based Medium in A Stirred Tank Reactor by A Ccpa Mutant of Lacticaseibacillus Caseilucia lopez lopezNo ratings yet
- Fermentation 04 00079Document9 pagesFermentation 04 00079CHIRANJEEVINo ratings yet
- Pone 0070516Document16 pagesPone 0070516Trân LâmNo ratings yet
- Jackson 2019 Artigo Resting CellsDocument10 pagesJackson 2019 Artigo Resting CellsCaio ToledoNo ratings yet
- s12934 017 0825 3Document16 pagess12934 017 0825 3Vashista KotraNo ratings yet
- Improved Production of Fatty Acid Ethyl...Document21 pagesImproved Production of Fatty Acid Ethyl...Maggie VenegasNo ratings yet
- Analysis of Zein by Matrix-Assisted Laser Desorption/Ionization Mass SpectrometryDocument6 pagesAnalysis of Zein by Matrix-Assisted Laser Desorption/Ionization Mass SpectrometrySabina BancilaNo ratings yet
- The Effect of Lignin Removal by Alkaline Peroxide Pretreatment On The Susceptibility of Corn Stover To Purified Cellulolytic and Xylanolytic EnzymesDocument11 pagesThe Effect of Lignin Removal by Alkaline Peroxide Pretreatment On The Susceptibility of Corn Stover To Purified Cellulolytic and Xylanolytic EnzymesAhmad Rama RamadhanNo ratings yet
- 2tolvanen, 2010, IJHE, Q-PCRDocument7 pages2tolvanen, 2010, IJHE, Q-PCRAdisNo ratings yet
- Morales AlvarezEDetal20173Document13 pagesMorales AlvarezEDetal20173Juan Diego CárdenasNo ratings yet
- Metabolic Capabilities of Actinobacillus Succinogenes For Succinic Acid ProductionDocument8 pagesMetabolic Capabilities of Actinobacillus Succinogenes For Succinic Acid Productionfrysye gumansalangiNo ratings yet
- Karbon Aktif, 1 (Naomi)Document12 pagesKarbon Aktif, 1 (Naomi)oktaNo ratings yet
- Lei 2001Document17 pagesLei 2001andres granadosNo ratings yet
- Biotechnological Strategies For Chitosan Production by Mucoralean Strains and Dimorphism Using Renewable SubstratesDocument15 pagesBiotechnological Strategies For Chitosan Production by Mucoralean Strains and Dimorphism Using Renewable SubstratesRoberta PippiNo ratings yet
- Zhang Et Al. - 2021 - Metabolism and Strategies For Enhanced Supply of ADocument14 pagesZhang Et Al. - 2021 - Metabolism and Strategies For Enhanced Supply of AAnaNo ratings yet
- 10 Publication 4Document10 pages10 Publication 4vdlu.hallNo ratings yet
- Schizochytrium Limacinum PDFDocument23 pagesSchizochytrium Limacinum PDFSheila CabreraNo ratings yet
- Energies: Second Generation Ethanol Production From Brewers' Spent GrainDocument12 pagesEnergies: Second Generation Ethanol Production From Brewers' Spent GrainmulerNo ratings yet
- Growth of E. Coli On Formate and Methanol. Nature Chemical BiologyDocument21 pagesGrowth of E. Coli On Formate and Methanol. Nature Chemical Biology3BGina FitrianingsihNo ratings yet
- P.L. Rogers, H.S. Shin and B. Wang - Biotransformation For L-Ephedrine ProductionDocument27 pagesP.L. Rogers, H.S. Shin and B. Wang - Biotransformation For L-Ephedrine ProductionRoundSTICNo ratings yet
- Influence of Chitosan Derivatization On PDFDocument10 pagesInfluence of Chitosan Derivatization On PDFOrlandoCialliNo ratings yet
- 2020 SILVA et al ( (6) -gingerol - β-cyclodextrin)Document10 pages2020 SILVA et al ( (6) -gingerol - β-cyclodextrin)Vitória ShiévilaNo ratings yet
- C5RA17797ADocument29 pagesC5RA17797AABB HisseinNo ratings yet
- (MB) Ortiz-Tena2016 Monosaccharides in CVDocument9 pages(MB) Ortiz-Tena2016 Monosaccharides in CVSacra PsyntergiaNo ratings yet
- Elagic AcidDocument13 pagesElagic Acidyousif husseinNo ratings yet
- Comparison of Methods For Detecting Protein Extracted From Excess Activated SludgeDocument17 pagesComparison of Methods For Detecting Protein Extracted From Excess Activated Sludgejean miguel oscorima celisNo ratings yet
- PRODUCCIÓN FUMARATO (1) GgyDocument7 pagesPRODUCCIÓN FUMARATO (1) GgyERICK GARCIA LAGUNANo ratings yet
- Biosinteză Microbiană Review 2018Document9 pagesBiosinteză Microbiană Review 2018Stas BicherschiNo ratings yet
- Water 11 00133Document13 pagesWater 11 00133yash sharmaNo ratings yet
- A Novel Recyclable Furoic Acid-Assisted Pretreatment For Sugarcane Bagasse Biorefinery in Co-Production of Xylooligosaccharides and GlucoseDocument8 pagesA Novel Recyclable Furoic Acid-Assisted Pretreatment For Sugarcane Bagasse Biorefinery in Co-Production of Xylooligosaccharides and GlucoseSthiden Sanabria ChaconNo ratings yet
- Advances in Microbial Bioconversion Mahto and GaraiDocument14 pagesAdvances in Microbial Bioconversion Mahto and GaraibanerjeetusharNo ratings yet
- A Novel Electroporation Procedure For Highly Efficient Transformation of Lipomyces StarkeyiDocument7 pagesA Novel Electroporation Procedure For Highly Efficient Transformation of Lipomyces Starkeyifilemon jaluNo ratings yet
- MS Ijbpas 2019 4888Document14 pagesMS Ijbpas 2019 4888leztlyNo ratings yet
- Energy Integration of Bioethanol Production Process From Algae Biomass: Comparison of SSF, SSCF and Acid HydrolysisDocument6 pagesEnergy Integration of Bioethanol Production Process From Algae Biomass: Comparison of SSF, SSCF and Acid HydrolysisDavinson Aguirre RodriguezNo ratings yet
- Red Seaweed BiorefineryDocument13 pagesRed Seaweed Biorefineryupright.hubNo ratings yet
- Optimization of B-Galactosidase Production From LaDocument9 pagesOptimization of B-Galactosidase Production From LaGaurav ChauhanNo ratings yet
- B-Galactosidases - A Reat Tool For Synthesizing Galactose-Containing CarbohydratesDocument15 pagesB-Galactosidases - A Reat Tool For Synthesizing Galactose-Containing CarbohydratesKukymovNo ratings yet
- Increased Isobutanol Production in Saccharomyces Cerevisiae by Eliminating Competing Pathways and Resolving Cofactor ImbalanceDocument12 pagesIncreased Isobutanol Production in Saccharomyces Cerevisiae by Eliminating Competing Pathways and Resolving Cofactor Imbalancejane doeNo ratings yet
- 2020 - Engineering E Coli LifespanDocument14 pages2020 - Engineering E Coli LifespanAkash AroraNo ratings yet
- Industrially Relevant Hydrolyzability and Fermentability of Sugarcane Bagasse Improved Effectively by Glycerol Organosolv PretreatmentDocument13 pagesIndustrially Relevant Hydrolyzability and Fermentability of Sugarcane Bagasse Improved Effectively by Glycerol Organosolv PretreatmentHarshad Vinay SavantNo ratings yet
- Fine Tuning of Nadh Oxidase Decreases Byproduct Accumulation in Respiration Deficient Xylose Metabolic Saccharomyces CerevisiaeDocument10 pagesFine Tuning of Nadh Oxidase Decreases Byproduct Accumulation in Respiration Deficient Xylose Metabolic Saccharomyces CerevisiaeideapadNo ratings yet
- Applied and Environmental Microbiology-2007-Schilling-499.fullDocument9 pagesApplied and Environmental Microbiology-2007-Schilling-499.fullchocosherNo ratings yet
- Modelling of Biogas Production From Glycerol by Anaerobic Process in A Baffled Multi-Stage DigestorDocument5 pagesModelling of Biogas Production From Glycerol by Anaerobic Process in A Baffled Multi-Stage DigestorEdgar Velastegui GonzálezNo ratings yet
- s13068 019 1604 7Document11 pagess13068 019 1604 7MARIA DE LA PAZ GRAJEDA PINEDANo ratings yet
- Microbial Sensing in FermentationFrom EverandMicrobial Sensing in FermentationSatinder K. BrarNo ratings yet
- Microbiological Studies On Dosa FermentationDocument9 pagesMicrobiological Studies On Dosa FermentationRit Wick100% (1)
- Edwin BarbosaDocument130 pagesEdwin BarbosaOscar GuillenNo ratings yet
- Boulardii 1Document11 pagesBoulardii 1janesprightNo ratings yet
- Saccharomyces Cerevisiae: Baker's YeastDocument7 pagesSaccharomyces Cerevisiae: Baker's YeastMicro ShamimNo ratings yet
- Studies On Bioethanol Production of Commercial Baker's and Alcohol Yeast Under Aerated Culture Using Sugarcane Molasses As The MediaDocument7 pagesStudies On Bioethanol Production of Commercial Baker's and Alcohol Yeast Under Aerated Culture Using Sugarcane Molasses As The MediaNabila AlyskaNo ratings yet
- Bellis Kullman. 2000. Application of Flow Cytometry For Measurement of Nuclear DNA Content in Fungi.Document16 pagesBellis Kullman. 2000. Application of Flow Cytometry For Measurement of Nuclear DNA Content in Fungi.Bellis KullmanNo ratings yet
- Yeast Beta Glucosidase Producer in WinemakingDocument13 pagesYeast Beta Glucosidase Producer in WinemakingSiti NurmilahNo ratings yet
- Ovber StartDocument12 pagesOvber StartHemali Sudhakar NarkhedeNo ratings yet
- Yepd AgarDocument2 pagesYepd AgarVeny Dwi NursandyNo ratings yet
- Microbial EnzymesDocument50 pagesMicrobial EnzymesJohnry Maloles100% (1)
- Ethanol Production From The Water Hyacinth Eichhornia Crassipes by Yeast Isolated From Various Hydro SpheresDocument4 pagesEthanol Production From The Water Hyacinth Eichhornia Crassipes by Yeast Isolated From Various Hydro SpheresmerinafitriaNo ratings yet
- Yeast Protocols: Third EditionDocument309 pagesYeast Protocols: Third Editionpavlina.mastrandreaNo ratings yet
- Bruce Alberts - Chapter1-8Document601 pagesBruce Alberts - Chapter1-8Tatheer Zahra100% (4)
- Brewing MicrobiologyDocument312 pagesBrewing MicrobiologyCristiane Moraes100% (1)
- (First Author) 2005 Journal-Of-BiotechnologyDocument189 pages(First Author) 2005 Journal-Of-BiotechnologyLicet Vanessa Durango CastillaNo ratings yet
- Chemical Engineering 414 Lab Manual 2013Document49 pagesChemical Engineering 414 Lab Manual 2013adamgarthNo ratings yet
- OutDocument12 pagesOutFiras Harish FadilaNo ratings yet
- Yeast - CIMADocument2 pagesYeast - CIMARichard RodriguezNo ratings yet
- Technical Data Sheet For Instant Dry Baker'S YeastDocument1 pageTechnical Data Sheet For Instant Dry Baker'S YeastRakesh SakarayNo ratings yet
- Characterization of Alcohol Resistant Yeast Saccharomyces Cerevisiae Isolated From ToddyDocument7 pagesCharacterization of Alcohol Resistant Yeast Saccharomyces Cerevisiae Isolated From ToddyfikerykNo ratings yet
- NPR 8 (4) 345-355Document11 pagesNPR 8 (4) 345-355Ricardo Telgie MoralesNo ratings yet
- Impact of Yeast-Handling Procedures On Beer Flavour Development During FermentationDocument7 pagesImpact of Yeast-Handling Procedures On Beer Flavour Development During FermentationMayra Schmidt RechsteinerNo ratings yet
- Wort Composition and Its Impact On The Flavour-Active Higher Alcohol and Ester Formation of Beer - A ReviewDocument7 pagesWort Composition and Its Impact On The Flavour-Active Higher Alcohol and Ester Formation of Beer - A ReviewGSBYGALATINo ratings yet
- Bioethanol ProductionDocument76 pagesBioethanol ProductionAsmamaw TesfawNo ratings yet
- YeastDocument43 pagesYeastResna N K Resi100% (1)
- Gin11 Counterselectable Marker PDFDocument10 pagesGin11 Counterselectable Marker PDFmadcaddietoo1600No ratings yet
- Dna RecombinantDocument580 pagesDna RecombinantSura100% (2)