
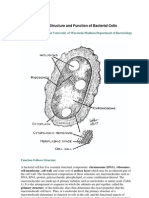
2016 Organisation and Control of Prokaryotic Genome
2016 Organisation and Control of Prokaryotic Genome
You might also like
- Khan Academy DNA Technology QuestionsDocument4 pagesKhan Academy DNA Technology QuestionsLoraNo ratings yet
- Molecular Parasitology 2016 PDFDocument546 pagesMolecular Parasitology 2016 PDFAura-Nicoleta Popinciuc0% (1)
- (H2) CI2.10 - Genetics of Bacteria (N) (STDT)Document37 pages(H2) CI2.10 - Genetics of Bacteria (N) (STDT)Timothy HandokoNo ratings yet
- Prokaryotic Genes - General Structures and Organization. Extrachromosomal Genome in Prokaryotes.Document2 pagesProkaryotic Genes - General Structures and Organization. Extrachromosomal Genome in Prokaryotes.carlottabovi28No ratings yet
- Prokaryotic and Eukaryotic CellsDocument122 pagesProkaryotic and Eukaryotic CellsAdithya NanuvalaNo ratings yet
- LECT 2 ProkaryotesDocument30 pagesLECT 2 ProkaryotesjudynakwebwaNo ratings yet
- A Guide To Wastewater MicrobiologyDocument14 pagesA Guide To Wastewater Microbiologyabduyassin8014No ratings yet
- Structure & Functions of Prokaryotic Cell. Bacteriophages.: Lecturer: - Sergei V. RedkozubovDocument54 pagesStructure & Functions of Prokaryotic Cell. Bacteriophages.: Lecturer: - Sergei V. RedkozubovJustineNo ratings yet
- Core Idea 2) Genetics and Inheritance: 2.9A) Structure and Organisation of Prokaryotic DnaDocument10 pagesCore Idea 2) Genetics and Inheritance: 2.9A) Structure and Organisation of Prokaryotic DnaReyes ChengNo ratings yet
- формы жизниDocument43 pagesформы жизниKhaled Mohammed GhalwashNo ratings yet
- StructureDocument15 pagesStructurefatimasanusimorikiNo ratings yet
- Mod 2Document85 pagesMod 2BikashGuptaNo ratings yet
- Full Microbiology With Diseases by Body System 4Th Edition Bauman Solutions Manual Online PDF All ChapterDocument32 pagesFull Microbiology With Diseases by Body System 4Th Edition Bauman Solutions Manual Online PDF All Chapterardeliatimor-leste23f100% (5)
- General Microbiology - Lecture 1Document41 pagesGeneral Microbiology - Lecture 1Almoatazbellah AbdallahNo ratings yet
- Bacterial ClassificationDocument5 pagesBacterial ClassificationShai KateNo ratings yet
- Week 11 Answers 2020Document8 pagesWeek 11 Answers 2020askldhfdasjkNo ratings yet
- Me 2 2Document47 pagesMe 2 2s93350190No ratings yet
- Infectious Disease NotesDocument43 pagesInfectious Disease NotessaguNo ratings yet
- Genetika Mikroba - Tugas 24 Nov 2016Document19 pagesGenetika Mikroba - Tugas 24 Nov 2016indrioctriNo ratings yet
- Bab 6 Sruktur Dan Fungsi Sel Bakteria (I)Document9 pagesBab 6 Sruktur Dan Fungsi Sel Bakteria (I)Shakira AmiraNo ratings yet
- Procaryotic Cell ArchitectureDocument18 pagesProcaryotic Cell ArchitectureWindi Dawn SallevaNo ratings yet
- Parker 2001Document6 pagesParker 2001Chinasa EkejiubaNo ratings yet
- Archean EonDocument8 pagesArchean EonJeremie LacroixNo ratings yet
- Module 1 GenomicsDocument19 pagesModule 1 GenomicsSiddhant MazumderNo ratings yet
- Unit 3 Medically Important MicroorganismsDocument9 pagesUnit 3 Medically Important Microorganisms2023800041No ratings yet
- Microbiology Lec 1 PDFDocument7 pagesMicrobiology Lec 1 PDFAhmed AliNo ratings yet
- Cell Biology Part 1Document29 pagesCell Biology Part 1hafiz.umer.7169No ratings yet
- Bt301 Midterm Subjectives 100% Guess Paper 2023 by Sulman AliDocument9 pagesBt301 Midterm Subjectives 100% Guess Paper 2023 by Sulman Aliahseera2005No ratings yet
- Components of PlasmidsDocument7 pagesComponents of PlasmidsKashifNo ratings yet
- E Tate@imperial Ac UkDocument19 pagesE Tate@imperial Ac UkSoRina SabauNo ratings yet
- Microbiology Summaries 1-30 Microbiology Summaries 1-30Document53 pagesMicrobiology Summaries 1-30 Microbiology Summaries 1-30yepNo ratings yet
- Assigment 1 BiologyDocument8 pagesAssigment 1 BiologyshimaabdollahpourNo ratings yet
- General Principles of Cell OrganisationDocument3 pagesGeneral Principles of Cell Organisation22194No ratings yet
- Functions of Organelles in CellDocument23 pagesFunctions of Organelles in CellZahid ShehzarNo ratings yet
- BACTERIADocument7 pagesBACTERIAsuruthi ramesh28No ratings yet
- FAQ in MicrobiologyDocument117 pagesFAQ in MicrobiologyVenkatesh Arumugam100% (2)
- Prokaryotic Cells Are Also CalledDocument25 pagesProkaryotic Cells Are Also CalledArdo RamdhaniNo ratings yet
- AP Biology Bacteria and VirusesDocument51 pagesAP Biology Bacteria and VirusesMichael TsaiNo ratings yet
- Bacteriology PDFDocument226 pagesBacteriology PDFWaseem Ur RahmanNo ratings yet
- Structure and Function of Cellular Genomes - Microbiology - OpenStaxDocument10 pagesStructure and Function of Cellular Genomes - Microbiology - OpenStaxAleksandra Sanja MartinovicNo ratings yet
- Micro 1 Chapter 1Document50 pagesMicro 1 Chapter 1chayiezen0301No ratings yet
- 4 1-4 2Document5 pages4 1-4 2owmnerdzNo ratings yet
- Abt PDFDocument9 pagesAbt PDFManikandan SundarapandiyanNo ratings yet
- Micro 1st ProkaryotesDocument16 pagesMicro 1st ProkaryotesMatahari SempurnaNo ratings yet
- SGL 2 (Structure of Bacterial Cell)Document32 pagesSGL 2 (Structure of Bacterial Cell)rekanshwan5No ratings yet
- Gene Organisation in Mito and ChloroplastDocument13 pagesGene Organisation in Mito and Chloroplastraryanraj44No ratings yet
- Mod 2Document24 pagesMod 2The GreatNo ratings yet
- Chromosomes, Nucleic Acids & Other Genetic ElementsDocument22 pagesChromosomes, Nucleic Acids & Other Genetic ElementsRadius JuliusNo ratings yet
- Genetic Material (F)Document40 pagesGenetic Material (F)PGMBslidesNo ratings yet
- Lecture 10 - ProkaryotesDocument64 pagesLecture 10 - Prokaryotesnm1006055No ratings yet
- Bacterial MorphologyDocument11 pagesBacterial Morphologydavid_physiologyNo ratings yet
- BIO206 NotesDocument7 pagesBIO206 NotesKaraLVNo ratings yet
- Act 7 Bacterial Morphology 1Document13 pagesAct 7 Bacterial Morphology 1Jez leah RamosNo ratings yet
- Fundamentals of MicrobiologyDocument51 pagesFundamentals of MicrobiologyJeul AzueloNo ratings yet
- BT 301 Short Question Mid TermDocument11 pagesBT 301 Short Question Mid TermAlishba khan100% (1)
- Eukaryotic Cell DefinitionDocument5 pagesEukaryotic Cell DefinitionAnonymous QtQewv0XQhNo ratings yet
- Chapter 4: Functional Anatomy of Prokaryotic and Eukaryotic CellsDocument94 pagesChapter 4: Functional Anatomy of Prokaryotic and Eukaryotic CellsTrevannie EdwardsNo ratings yet
- Bacterial Cell Structure: BacteriaDocument5 pagesBacterial Cell Structure: BacteriaHassan AljaberiNo ratings yet
- CH-3. Microbial Cell StructureDocument13 pagesCH-3. Microbial Cell StructureQur' anNo ratings yet
- Form 4Hk/Year 10 Biology Characteristics and Diagrams of Bacteria, Viruses and FungiDocument12 pagesForm 4Hk/Year 10 Biology Characteristics and Diagrams of Bacteria, Viruses and FungiShadreck MagangaNo ratings yet
- Biology Unleashed: A Comprehensive Guide to Mastering the Science of LifeFrom EverandBiology Unleashed: A Comprehensive Guide to Mastering the Science of LifeNo ratings yet
- The First Comparative Examination 2021-22: Class XII (ISC) BIOLOGY Paper-1 (Theory)Document5 pagesThe First Comparative Examination 2021-22: Class XII (ISC) BIOLOGY Paper-1 (Theory)Chinmay SinghNo ratings yet
- 08 Genes and MutationDocument16 pages08 Genes and Mutationapi-255000301No ratings yet
- Randomized Biobrick Assembly: A Novel Dna Assembly Method For Randomizing and Optimizing Genetic Circuits and Metabolic PathwaysDocument25 pagesRandomized Biobrick Assembly: A Novel Dna Assembly Method For Randomizing and Optimizing Genetic Circuits and Metabolic PathwaysShampa SenNo ratings yet
- 12 Fire Letters - 1 DNA Strand Template (1 Fire Letter Sequence 144 Vector Codes)Document1 page12 Fire Letters - 1 DNA Strand Template (1 Fire Letter Sequence 144 Vector Codes)RobertNo ratings yet
- Quarter 2 Week 4 Genetic Engineering 1Document15 pagesQuarter 2 Week 4 Genetic Engineering 1Sedrick Kyle BautistaNo ratings yet
- Yeast Vector (PYJ)Document24 pagesYeast Vector (PYJ)Shabeer Ahmad0% (2)
- MeiosisDocument3 pagesMeiosisLaasya ChitrakshiNo ratings yet
- Feschotte 2007Document38 pagesFeschotte 2007Axel GómezNo ratings yet
- Genetic Engineering: Unit 3Document60 pagesGenetic Engineering: Unit 3blackmoneygrabberNo ratings yet
- Chromatin RemodellingDocument234 pagesChromatin Remodellingplastioid4079No ratings yet
- 2 Fillable PDF-Meiosis WebQuest Student HandoutDocument5 pages2 Fillable PDF-Meiosis WebQuest Student HandoutAngelNo ratings yet
- Tesis doctoral de Bryan Keith Holland - “Discovery of mature microRNA sequences within the protein-coding regions of global HIV-1 genomes - Predictions of novel mechanisms for viral infection and pathogenicity”Document183 pagesTesis doctoral de Bryan Keith Holland - “Discovery of mature microRNA sequences within the protein-coding regions of global HIV-1 genomes - Predictions of novel mechanisms for viral infection and pathogenicity”Omar Alejandro Herrera ArenasNo ratings yet
- The Complete Genome Sequence of The Gram-Positive Bacterium Bacillus SubtilisDocument18 pagesThe Complete Genome Sequence of The Gram-Positive Bacterium Bacillus SubtilisPaco BaGaNo ratings yet
- Module 11Document64 pagesModule 11Andrew Dela CruzNo ratings yet
- 2nd Sem Q1 Week 1 Lesson 1 Genetic EngineeringDocument42 pages2nd Sem Q1 Week 1 Lesson 1 Genetic Engineeringayesha iris matillaNo ratings yet
- II TranscriptionDocument46 pagesII TranscriptionGail IbanezNo ratings yet
- Harper S Illustrated Biochemistry by Vic-385-403Document19 pagesHarper S Illustrated Biochemistry by Vic-385-403DavidNo ratings yet
- Answerkey ExamI 13Document10 pagesAnswerkey ExamI 13waldo625No ratings yet
- (H2) CI2.4 - Gene MutationDocument17 pages(H2) CI2.4 - Gene MutationTimothy HandokoNo ratings yet
- N 5 DZNJ2 P BO0 Ukp BZDocument11 pagesN 5 DZNJ2 P BO0 Ukp BZBackup IsaNo ratings yet
- Molecular Basis of InheritanceDocument3 pagesMolecular Basis of Inheritancesamarpeetnandanwar21No ratings yet
- Performance Task in ELSDocument16 pagesPerformance Task in ELSMaria Kristina BernadetteNo ratings yet
- Cloning Tech GuideDocument40 pagesCloning Tech GuideMia C.100% (1)
- Bio Ninja Unit 7 MergedDocument4 pagesBio Ninja Unit 7 Mergedshwooby shawoobyNo ratings yet
- Fidelity of DNA Replication: John D. Roberts' and Thomas A. Kunke12Document31 pagesFidelity of DNA Replication: John D. Roberts' and Thomas A. Kunke12Kavi GourNo ratings yet
- Integration Vectors For Gram PossitiveDocument58 pagesIntegration Vectors For Gram PossitiveQuynh Anh Nguyen100% (1)
- PCGPV VectorDocument1 pagePCGPV VectorAlleleBiotechNo ratings yet
- 10.06 Prokaryotic TranslationDocument4 pages10.06 Prokaryotic TranslationWidya NingsiNo ratings yet

























































































Download as doc, pdf, or txt
You might also like
- Khan Academy DNA Technology QuestionsDocument4 pagesKhan Academy DNA Technology QuestionsLoraNo ratings yet
- Molecular Parasitology 2016 PDFDocument546 pagesMolecular Parasitology 2016 PDFAura-Nicoleta Popinciuc0% (1)
- (H2) CI2.10 - Genetics of Bacteria (N) (STDT)Document37 pages(H2) CI2.10 - Genetics of Bacteria (N) (STDT)Timothy HandokoNo ratings yet
- Prokaryotic Genes - General Structures and Organization. Extrachromosomal Genome in Prokaryotes.Document2 pagesProkaryotic Genes - General Structures and Organization. Extrachromosomal Genome in Prokaryotes.carlottabovi28No ratings yet
- Prokaryotic and Eukaryotic CellsDocument122 pagesProkaryotic and Eukaryotic CellsAdithya NanuvalaNo ratings yet
- LECT 2 ProkaryotesDocument30 pagesLECT 2 ProkaryotesjudynakwebwaNo ratings yet
- A Guide To Wastewater MicrobiologyDocument14 pagesA Guide To Wastewater Microbiologyabduyassin8014No ratings yet
- Structure & Functions of Prokaryotic Cell. Bacteriophages.: Lecturer: - Sergei V. RedkozubovDocument54 pagesStructure & Functions of Prokaryotic Cell. Bacteriophages.: Lecturer: - Sergei V. RedkozubovJustineNo ratings yet
- Core Idea 2) Genetics and Inheritance: 2.9A) Structure and Organisation of Prokaryotic DnaDocument10 pagesCore Idea 2) Genetics and Inheritance: 2.9A) Structure and Organisation of Prokaryotic DnaReyes ChengNo ratings yet
- формы жизниDocument43 pagesформы жизниKhaled Mohammed GhalwashNo ratings yet
- StructureDocument15 pagesStructurefatimasanusimorikiNo ratings yet
- Mod 2Document85 pagesMod 2BikashGuptaNo ratings yet
- Full Microbiology With Diseases by Body System 4Th Edition Bauman Solutions Manual Online PDF All ChapterDocument32 pagesFull Microbiology With Diseases by Body System 4Th Edition Bauman Solutions Manual Online PDF All Chapterardeliatimor-leste23f100% (5)
- General Microbiology - Lecture 1Document41 pagesGeneral Microbiology - Lecture 1Almoatazbellah AbdallahNo ratings yet
- Bacterial ClassificationDocument5 pagesBacterial ClassificationShai KateNo ratings yet
- Week 11 Answers 2020Document8 pagesWeek 11 Answers 2020askldhfdasjkNo ratings yet
- Me 2 2Document47 pagesMe 2 2s93350190No ratings yet
- Infectious Disease NotesDocument43 pagesInfectious Disease NotessaguNo ratings yet
- Genetika Mikroba - Tugas 24 Nov 2016Document19 pagesGenetika Mikroba - Tugas 24 Nov 2016indrioctriNo ratings yet
- Bab 6 Sruktur Dan Fungsi Sel Bakteria (I)Document9 pagesBab 6 Sruktur Dan Fungsi Sel Bakteria (I)Shakira AmiraNo ratings yet
- Procaryotic Cell ArchitectureDocument18 pagesProcaryotic Cell ArchitectureWindi Dawn SallevaNo ratings yet
- Parker 2001Document6 pagesParker 2001Chinasa EkejiubaNo ratings yet
- Archean EonDocument8 pagesArchean EonJeremie LacroixNo ratings yet
- Module 1 GenomicsDocument19 pagesModule 1 GenomicsSiddhant MazumderNo ratings yet
- Unit 3 Medically Important MicroorganismsDocument9 pagesUnit 3 Medically Important Microorganisms2023800041No ratings yet
- Microbiology Lec 1 PDFDocument7 pagesMicrobiology Lec 1 PDFAhmed AliNo ratings yet
- Cell Biology Part 1Document29 pagesCell Biology Part 1hafiz.umer.7169No ratings yet
- Bt301 Midterm Subjectives 100% Guess Paper 2023 by Sulman AliDocument9 pagesBt301 Midterm Subjectives 100% Guess Paper 2023 by Sulman Aliahseera2005No ratings yet
- Components of PlasmidsDocument7 pagesComponents of PlasmidsKashifNo ratings yet
- E Tate@imperial Ac UkDocument19 pagesE Tate@imperial Ac UkSoRina SabauNo ratings yet
- Microbiology Summaries 1-30 Microbiology Summaries 1-30Document53 pagesMicrobiology Summaries 1-30 Microbiology Summaries 1-30yepNo ratings yet
- Assigment 1 BiologyDocument8 pagesAssigment 1 BiologyshimaabdollahpourNo ratings yet
- General Principles of Cell OrganisationDocument3 pagesGeneral Principles of Cell Organisation22194No ratings yet
- Functions of Organelles in CellDocument23 pagesFunctions of Organelles in CellZahid ShehzarNo ratings yet
- BACTERIADocument7 pagesBACTERIAsuruthi ramesh28No ratings yet
- FAQ in MicrobiologyDocument117 pagesFAQ in MicrobiologyVenkatesh Arumugam100% (2)
- Prokaryotic Cells Are Also CalledDocument25 pagesProkaryotic Cells Are Also CalledArdo RamdhaniNo ratings yet
- AP Biology Bacteria and VirusesDocument51 pagesAP Biology Bacteria and VirusesMichael TsaiNo ratings yet
- Bacteriology PDFDocument226 pagesBacteriology PDFWaseem Ur RahmanNo ratings yet
- Structure and Function of Cellular Genomes - Microbiology - OpenStaxDocument10 pagesStructure and Function of Cellular Genomes - Microbiology - OpenStaxAleksandra Sanja MartinovicNo ratings yet
- Micro 1 Chapter 1Document50 pagesMicro 1 Chapter 1chayiezen0301No ratings yet
- 4 1-4 2Document5 pages4 1-4 2owmnerdzNo ratings yet
- Abt PDFDocument9 pagesAbt PDFManikandan SundarapandiyanNo ratings yet
- Micro 1st ProkaryotesDocument16 pagesMicro 1st ProkaryotesMatahari SempurnaNo ratings yet
- SGL 2 (Structure of Bacterial Cell)Document32 pagesSGL 2 (Structure of Bacterial Cell)rekanshwan5No ratings yet
- Gene Organisation in Mito and ChloroplastDocument13 pagesGene Organisation in Mito and Chloroplastraryanraj44No ratings yet
- Mod 2Document24 pagesMod 2The GreatNo ratings yet
- Chromosomes, Nucleic Acids & Other Genetic ElementsDocument22 pagesChromosomes, Nucleic Acids & Other Genetic ElementsRadius JuliusNo ratings yet
- Genetic Material (F)Document40 pagesGenetic Material (F)PGMBslidesNo ratings yet
- Lecture 10 - ProkaryotesDocument64 pagesLecture 10 - Prokaryotesnm1006055No ratings yet
- Bacterial MorphologyDocument11 pagesBacterial Morphologydavid_physiologyNo ratings yet
- BIO206 NotesDocument7 pagesBIO206 NotesKaraLVNo ratings yet
- Act 7 Bacterial Morphology 1Document13 pagesAct 7 Bacterial Morphology 1Jez leah RamosNo ratings yet
- Fundamentals of MicrobiologyDocument51 pagesFundamentals of MicrobiologyJeul AzueloNo ratings yet
- BT 301 Short Question Mid TermDocument11 pagesBT 301 Short Question Mid TermAlishba khan100% (1)
- Eukaryotic Cell DefinitionDocument5 pagesEukaryotic Cell DefinitionAnonymous QtQewv0XQhNo ratings yet
- Chapter 4: Functional Anatomy of Prokaryotic and Eukaryotic CellsDocument94 pagesChapter 4: Functional Anatomy of Prokaryotic and Eukaryotic CellsTrevannie EdwardsNo ratings yet
- Bacterial Cell Structure: BacteriaDocument5 pagesBacterial Cell Structure: BacteriaHassan AljaberiNo ratings yet
- CH-3. Microbial Cell StructureDocument13 pagesCH-3. Microbial Cell StructureQur' anNo ratings yet
- Form 4Hk/Year 10 Biology Characteristics and Diagrams of Bacteria, Viruses and FungiDocument12 pagesForm 4Hk/Year 10 Biology Characteristics and Diagrams of Bacteria, Viruses and FungiShadreck MagangaNo ratings yet
- Biology Unleashed: A Comprehensive Guide to Mastering the Science of LifeFrom EverandBiology Unleashed: A Comprehensive Guide to Mastering the Science of LifeNo ratings yet
- The First Comparative Examination 2021-22: Class XII (ISC) BIOLOGY Paper-1 (Theory)Document5 pagesThe First Comparative Examination 2021-22: Class XII (ISC) BIOLOGY Paper-1 (Theory)Chinmay SinghNo ratings yet
- 08 Genes and MutationDocument16 pages08 Genes and Mutationapi-255000301No ratings yet
- Randomized Biobrick Assembly: A Novel Dna Assembly Method For Randomizing and Optimizing Genetic Circuits and Metabolic PathwaysDocument25 pagesRandomized Biobrick Assembly: A Novel Dna Assembly Method For Randomizing and Optimizing Genetic Circuits and Metabolic PathwaysShampa SenNo ratings yet
- 12 Fire Letters - 1 DNA Strand Template (1 Fire Letter Sequence 144 Vector Codes)Document1 page12 Fire Letters - 1 DNA Strand Template (1 Fire Letter Sequence 144 Vector Codes)RobertNo ratings yet
- Quarter 2 Week 4 Genetic Engineering 1Document15 pagesQuarter 2 Week 4 Genetic Engineering 1Sedrick Kyle BautistaNo ratings yet
- Yeast Vector (PYJ)Document24 pagesYeast Vector (PYJ)Shabeer Ahmad0% (2)
- MeiosisDocument3 pagesMeiosisLaasya ChitrakshiNo ratings yet
- Feschotte 2007Document38 pagesFeschotte 2007Axel GómezNo ratings yet
- Genetic Engineering: Unit 3Document60 pagesGenetic Engineering: Unit 3blackmoneygrabberNo ratings yet
- Chromatin RemodellingDocument234 pagesChromatin Remodellingplastioid4079No ratings yet
- 2 Fillable PDF-Meiosis WebQuest Student HandoutDocument5 pages2 Fillable PDF-Meiosis WebQuest Student HandoutAngelNo ratings yet
- Tesis doctoral de Bryan Keith Holland - “Discovery of mature microRNA sequences within the protein-coding regions of global HIV-1 genomes - Predictions of novel mechanisms for viral infection and pathogenicity”Document183 pagesTesis doctoral de Bryan Keith Holland - “Discovery of mature microRNA sequences within the protein-coding regions of global HIV-1 genomes - Predictions of novel mechanisms for viral infection and pathogenicity”Omar Alejandro Herrera ArenasNo ratings yet
- The Complete Genome Sequence of The Gram-Positive Bacterium Bacillus SubtilisDocument18 pagesThe Complete Genome Sequence of The Gram-Positive Bacterium Bacillus SubtilisPaco BaGaNo ratings yet
- Module 11Document64 pagesModule 11Andrew Dela CruzNo ratings yet
- 2nd Sem Q1 Week 1 Lesson 1 Genetic EngineeringDocument42 pages2nd Sem Q1 Week 1 Lesson 1 Genetic Engineeringayesha iris matillaNo ratings yet
- II TranscriptionDocument46 pagesII TranscriptionGail IbanezNo ratings yet
- Harper S Illustrated Biochemistry by Vic-385-403Document19 pagesHarper S Illustrated Biochemistry by Vic-385-403DavidNo ratings yet
- Answerkey ExamI 13Document10 pagesAnswerkey ExamI 13waldo625No ratings yet
- (H2) CI2.4 - Gene MutationDocument17 pages(H2) CI2.4 - Gene MutationTimothy HandokoNo ratings yet
- N 5 DZNJ2 P BO0 Ukp BZDocument11 pagesN 5 DZNJ2 P BO0 Ukp BZBackup IsaNo ratings yet
- Molecular Basis of InheritanceDocument3 pagesMolecular Basis of Inheritancesamarpeetnandanwar21No ratings yet
- Performance Task in ELSDocument16 pagesPerformance Task in ELSMaria Kristina BernadetteNo ratings yet
- Cloning Tech GuideDocument40 pagesCloning Tech GuideMia C.100% (1)
- Bio Ninja Unit 7 MergedDocument4 pagesBio Ninja Unit 7 Mergedshwooby shawoobyNo ratings yet
- Fidelity of DNA Replication: John D. Roberts' and Thomas A. Kunke12Document31 pagesFidelity of DNA Replication: John D. Roberts' and Thomas A. Kunke12Kavi GourNo ratings yet
- Integration Vectors For Gram PossitiveDocument58 pagesIntegration Vectors For Gram PossitiveQuynh Anh Nguyen100% (1)
- PCGPV VectorDocument1 pagePCGPV VectorAlleleBiotechNo ratings yet
- 10.06 Prokaryotic TranslationDocument4 pages10.06 Prokaryotic TranslationWidya NingsiNo ratings yet