
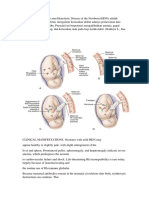
Trias Et Al.
Trias Et Al.
You might also like
- Hand Hygiene Knowledge Assessment Questionnaire (IHI)Document2 pagesHand Hygiene Knowledge Assessment Questionnaire (IHI)Suseno Aji67% (3)
- Basic Concepts On Laboratory Biosafety and BiosecurityDocument7 pagesBasic Concepts On Laboratory Biosafety and BiosecurityKristine PangahinNo ratings yet
- The Immune SystemDocument19 pagesThe Immune Systemmay esteban100% (2)
- ajol-file-journals_82_articles_82639_submission_proof_82639-973-198882-1-10-20121025Document8 pagesajol-file-journals_82_articles_82639_submission_proof_82639-973-198882-1-10-20121025yarach9494No ratings yet
- Food ControlDocument7 pagesFood ControlValentin GhenceaNo ratings yet
- Officejmbfs,+Jmbfs 1626 OyusaglamDocument5 pagesOfficejmbfs,+Jmbfs 1626 OyusaglamWendy SarmientoNo ratings yet
- Food Microbiology 85 (2020) 103282Document8 pagesFood Microbiology 85 (2020) 103282shinchanNo ratings yet
- Example Article IJFMDocument12 pagesExample Article IJFMMarcel HoungbédjiNo ratings yet
- Microbiological assessment of fresh, minimally processed vegetables from open air markets and supermarkets in Luzon, Philippines, for food safetyDocument10 pagesMicrobiological assessment of fresh, minimally processed vegetables from open air markets and supermarkets in Luzon, Philippines, for food safetyiyaNo ratings yet
- 1 Pilihan Hari JugaDocument9 pages1 Pilihan Hari JuganadyasuwayviaNo ratings yet
- Fresh Cut Pineapple As A New Carrier of Probiotic Lactic Acid BacteriaDocument10 pagesFresh Cut Pineapple As A New Carrier of Probiotic Lactic Acid Bacteriahossein alirezaeiNo ratings yet
- Armando Et Al 2011Document10 pagesArmando Et Al 2011Adrian Melgratti JobsonNo ratings yet
- 2018 - Adebayo & Martínez-Carrera - IJ Food Science & TechnologyDocument15 pages2018 - Adebayo & Martínez-Carrera - IJ Food Science & TechnologyAdair Abundis IbarraNo ratings yet
- 90Document11 pages90Eka Nurul HayatiNo ratings yet
- Biological Control: Min Zheng, Jingying Shi, Jian Shi, Qingguo Wang, Yanhua LiDocument7 pagesBiological Control: Min Zheng, Jingying Shi, Jian Shi, Qingguo Wang, Yanhua LiKieu Viet AnhNo ratings yet
- Klaus 2021Document14 pagesKlaus 2021Juan QuitianNo ratings yet
- 7IJEEFUSAUG20197Document8 pages7IJEEFUSAUG20197TJPRC PublicationsNo ratings yet
- C0 Formulir PemohonanDocument5 pagesC0 Formulir PemohonanDiah AyuningrumNo ratings yet
- Escherichia Coli O157:H7, and Listeria Monocytogenes On Fresh LettuceDocument6 pagesEscherichia Coli O157:H7, and Listeria Monocytogenes On Fresh LettuceTanmoy Kumar DeyNo ratings yet
- Antibacterial Mechanism of Lactic Acid On Physiological andDocument6 pagesAntibacterial Mechanism of Lactic Acid On Physiological andlox standardNo ratings yet
- 1 s2.0 S0362028X22084447 MainDocument9 pages1 s2.0 S0362028X22084447 MainTanti 01No ratings yet
- TocoshDocument8 pagesTocoshHansNo ratings yet
- 2021 Antagonistic Activity of Lactic Acid Bacteria Against Phytopathogenic Fungi Isolated From Cherry TomatoDocument10 pages2021 Antagonistic Activity of Lactic Acid Bacteria Against Phytopathogenic Fungi Isolated From Cherry TomatoEduardo Urbano Moraga CáceresNo ratings yet
- Furlaneto-Maia2020 Article AntimicrobialActivityOfEnterocDocument10 pagesFurlaneto-Maia2020 Article AntimicrobialActivityOfEnterocAmanuel AdugnaNo ratings yet
- 45 fr-2021-446 AliDocument9 pages45 fr-2021-446 AliHikmah KimaNo ratings yet
- Isolation of Antibacterial Components From Infusion of Caesalpinia ParaguariensisDocument10 pagesIsolation of Antibacterial Components From Infusion of Caesalpinia ParaguariensisAndres Fernando Silvestre SuarezNo ratings yet
- Nutrients 09 00755Document22 pagesNutrients 09 00755John James MonteverdeNo ratings yet
- Food Microbiology: Meenu N. Perera, Tamar Abuladze, Manrong Li, Joelle Woolston, Alexander SulakvelidzeDocument7 pagesFood Microbiology: Meenu N. Perera, Tamar Abuladze, Manrong Li, Joelle Woolston, Alexander Sulakvelidzeapria siskaNo ratings yet
- The Chemical Preservatives Used To ControlDocument6 pagesThe Chemical Preservatives Used To ControlAmine BenineNo ratings yet
- Food Chemistry: Analytical MethodsDocument5 pagesFood Chemistry: Analytical MethodsFatehatun NoorNo ratings yet
- Reuteri 2-20B and Pediococcus Acidilactici 0-11A Isolated From FuraDocument9 pagesReuteri 2-20B and Pediococcus Acidilactici 0-11A Isolated From FuramostecocNo ratings yet
- 1 s2.0 S0362028X22018543 MainDocument4 pages1 s2.0 S0362028X22018543 MainVNo ratings yet
- Probiotics As Control Agents in AquacultureDocument4 pagesProbiotics As Control Agents in AquacultureBerlin Coldfish BarkerNo ratings yet
- Lactobacillus BrevisDocument21 pagesLactobacillus Brevisshreya singhNo ratings yet
- Pan, X., Chen, F., Wu, T., Tang, H., Dan Zhao, Z. 2009. The Acid, Bile Tolerance and Antimicrobial Property of Lactobacillus Acidophilus NIT. J. Food Control. 20 598-602.Document5 pagesPan, X., Chen, F., Wu, T., Tang, H., Dan Zhao, Z. 2009. The Acid, Bile Tolerance and Antimicrobial Property of Lactobacillus Acidophilus NIT. J. Food Control. 20 598-602.AfdhalRuslanNo ratings yet
- Research Article: in Vitro Evaluation of Probiotic Properties of Lactic Acid BacteriaDocument12 pagesResearch Article: in Vitro Evaluation of Probiotic Properties of Lactic Acid BacteriaNatalija Atanasova-PancevskaNo ratings yet
- Budu BiotekDocument5 pagesBudu BiotekIndra Yusuf PratamaNo ratings yet
- BacteriophagesDocument5 pagesBacteriophagesJames UgbesNo ratings yet
- Characterisation of Bacteriocins Produced by LactoDocument10 pagesCharacterisation of Bacteriocins Produced by Lactokhurry testNo ratings yet
- Franco 2016Document15 pagesFranco 2016RogerioportelaNo ratings yet
- Bacteriocins From Lactic Acid Bacteria Inhibit Food Borne Pathogens.Document7 pagesBacteriocins From Lactic Acid Bacteria Inhibit Food Borne Pathogens.International Organization of Scientific Research (IOSR)No ratings yet
- Invitro Probiotic Evaluation of Lactic Acid BacteriaDocument8 pagesInvitro Probiotic Evaluation of Lactic Acid Bacteria325071067No ratings yet
- EFFECTS OF CALCIUM ASCORBATE AND CALCIUM LACTATE ON QUALITY OF FRESH-CUT PINEAPPLE (Ananas Comosus) (#544974) - 709246Document8 pagesEFFECTS OF CALCIUM ASCORBATE AND CALCIUM LACTATE ON QUALITY OF FRESH-CUT PINEAPPLE (Ananas Comosus) (#544974) - 709246Gaganpreet KaurNo ratings yet
- Journal RosettoDocument9 pagesJournal RosettoBrad PorneaNo ratings yet
- Food Control: Djamel Djenane, Javier Yangüela, Luis Montañés, Mouloud Djerbal, Pedro RoncalésDocument8 pagesFood Control: Djamel Djenane, Javier Yangüela, Luis Montañés, Mouloud Djerbal, Pedro RoncalésEstefaníaNo ratings yet
- Abo-State Et Al., 2015Document7 pagesAbo-State Et Al., 2015Jordana KalineNo ratings yet
- Martínez Et Al 2018Document10 pagesMartínez Et Al 2018Adrian Melgratti JobsonNo ratings yet
- In-Vitro Screening of Antibacterial Activity of Lactic Acid Bacteria Against Common Enteric PathogensDocument6 pagesIn-Vitro Screening of Antibacterial Activity of Lactic Acid Bacteria Against Common Enteric PathogensInternational Medical PublisherNo ratings yet
- Acid LaticoDocument9 pagesAcid LaticoWilson SilvaNo ratings yet
- The Lactic Acid Bacteria (Lab) Activities Against Pathogenic and Spoilage Bacteria On A Dried Salted LoligoDocument8 pagesThe Lactic Acid Bacteria (Lab) Activities Against Pathogenic and Spoilage Bacteria On A Dried Salted LoligocumabuatdownloadNo ratings yet
- In Vitro SourdoughDocument11 pagesIn Vitro SourdoughMaryam JameelahNo ratings yet
- EuglenaDocument8 pagesEuglenaHAROLD HUMBERTO REYES ALVAREZNo ratings yet
- (Allameh, 2012) Isolation, Identification and Characterization of Leuconostoc Mesenteroides As A New Probiotic From Intestine of Snakehead Fish (Channa Striatus) .Document7 pages(Allameh, 2012) Isolation, Identification and Characterization of Leuconostoc Mesenteroides As A New Probiotic From Intestine of Snakehead Fish (Channa Striatus) .José Luis Plaza DoradoNo ratings yet
- Barbosa2013 PDFDocument6 pagesBarbosa2013 PDFSilvia Guerrero GonzalezNo ratings yet
- Holz LAB AntifungalDocument17 pagesHolz LAB AntifungalIstván NagyNo ratings yet
- Ochratoxin A Induced Growth Disturbance, Immune Suppression, and Histopathological Changes of Liver and Kidney of Nil...Document18 pagesOchratoxin A Induced Growth Disturbance, Immune Suppression, and Histopathological Changes of Liver and Kidney of Nil...Abdallah Tag Eldein MansourNo ratings yet
- Safe Practices for Food Processes _ Analysis and Evaluation of Preventive Control Measures for the Control and Reduction_Elimination of Microbial Hazards on Fresh and Fresh-Cut Produce_ List of ReferencesDocument41 pagesSafe Practices for Food Processes _ Analysis and Evaluation of Preventive Control Measures for the Control and Reduction_Elimination of Microbial Hazards on Fresh and Fresh-Cut Produce_ List of ReferencesMohamed ElsawyNo ratings yet
- MicrobiologicalDocument12 pagesMicrobiologicalTrung Truong NhatNo ratings yet
- Preliminary Characterization of BacterioDocument9 pagesPreliminary Characterization of Bacteriomanuomar35No ratings yet
- BacteriocinDocument16 pagesBacteriocintransformers7No ratings yet
- Antioxidant and Antibacterial Activities of Polyphenols From Ethnomedicinal Plants of Burkina FasoDocument6 pagesAntioxidant and Antibacterial Activities of Polyphenols From Ethnomedicinal Plants of Burkina Fasokaori_lawlietNo ratings yet
- Antimicrobial Activities of An Oxygenated Cyclohexanone Derivative Isolated FromDocument36 pagesAntimicrobial Activities of An Oxygenated Cyclohexanone Derivative Isolated FromUtari ZulfianiNo ratings yet
- Nursing Care For A Family Caring For Children With Infectious DiseaseDocument4 pagesNursing Care For A Family Caring For Children With Infectious DiseasekdfhjfhfNo ratings yet
- Endocrine NotesDocument157 pagesEndocrine NotesLaila Cristine CarvalhoNo ratings yet
- Bacteriology Mtap (Part 2)Document10 pagesBacteriology Mtap (Part 2)Joshua CayagoNo ratings yet
- Bacon 2006Document10 pagesBacon 2006Bryan Lloyd LauritoNo ratings yet
- Histology Lab GuideDocument96 pagesHistology Lab GuidevrindaNo ratings yet
- HPV16 Genetic Variation and The Development of Cervical Cancer WorldwideDocument5 pagesHPV16 Genetic Variation and The Development of Cervical Cancer WorldwideQuyết Lê HữuNo ratings yet
- General Biology 2Document5 pagesGeneral Biology 2Kim Ashley Padernal MandronNo ratings yet
- Signaling of Receptor Tyrosine Kinases in The Nucleus: Sally-Anne Stephenson, Inga Mertens-Walker and Adrian HeringtonDocument25 pagesSignaling of Receptor Tyrosine Kinases in The Nucleus: Sally-Anne Stephenson, Inga Mertens-Walker and Adrian HeringtonAyu SafitriNo ratings yet
- Current UIP (Universal Immunization Programme) Schedule in Our StateDocument35 pagesCurrent UIP (Universal Immunization Programme) Schedule in Our StateGopireddy Sivakarthik ReddyNo ratings yet
- Antibiotic ProductionDocument19 pagesAntibiotic ProductionBright OgwoNo ratings yet
- HDN Alias Eritroblastosis FetalisDocument6 pagesHDN Alias Eritroblastosis FetalisOlivia RamadhaniNo ratings yet
- Biological Classification MCQDocument17 pagesBiological Classification MCQNawaraj RokayaNo ratings yet
- Science and Technology Sep June EnglishDocument85 pagesScience and Technology Sep June EnglishJAGADEESH AMNo ratings yet
- CeftolozaneTazobactam A New Option in The TreatmenDocument7 pagesCeftolozaneTazobactam A New Option in The TreatmenAisyah RohieNo ratings yet
- Gene Action and Non-Mendelian InheritanceDocument39 pagesGene Action and Non-Mendelian InheritanceFrank AyrosoNo ratings yet
- Chapter 10: Muscle Tissue: VoluntaryDocument13 pagesChapter 10: Muscle Tissue: VoluntaryClyde BaltazarNo ratings yet
- Effectofberberineon GiardiasisDocument5 pagesEffectofberberineon GiardiasisSummiNo ratings yet
- Quiz RenalDocument81 pagesQuiz RenalMedShare86% (14)
- Avanced Lipoproteins Reserch 2017Document108 pagesAvanced Lipoproteins Reserch 2017Carlos SorianoNo ratings yet
- Arroz Con Mango PDFDocument197 pagesArroz Con Mango PDFVanshika JainNo ratings yet
- HBs Ag Q2Document8 pagesHBs Ag Q2Naveen Kumar MNo ratings yet
- Mutation: Altered Genes: Apago PDF EnhancerDocument5 pagesMutation: Altered Genes: Apago PDF EnhancerAndresNo ratings yet
- The Buzz On Caffeine in Invertebrates Effects On Behavior and Molecular MechanismsDocument9 pagesThe Buzz On Caffeine in Invertebrates Effects On Behavior and Molecular Mechanismsittoiram setagNo ratings yet
- Conceptual Issues in Patenting of Life Forms.Document7 pagesConceptual Issues in Patenting of Life Forms.Jay PatelNo ratings yet
- Antifungal Azole Derivatives and Their Pharmacological Potential: Prospects & RetrospectsDocument14 pagesAntifungal Azole Derivatives and Their Pharmacological Potential: Prospects & RetrospectsRenata YolandaNo ratings yet
- Tetracycline's: Jagir R. Patel Asst Prof Dept. Pharmacology Anand Pharmacy CollegeDocument21 pagesTetracycline's: Jagir R. Patel Asst Prof Dept. Pharmacology Anand Pharmacy CollegeJagirNo ratings yet
- Master Thesis Cell CultureDocument5 pagesMaster Thesis Cell Cultureebonybatesshreveport100% (2)
























































































Download as pdf or txt
You might also like
- Hand Hygiene Knowledge Assessment Questionnaire (IHI)Document2 pagesHand Hygiene Knowledge Assessment Questionnaire (IHI)Suseno Aji67% (3)
- Basic Concepts On Laboratory Biosafety and BiosecurityDocument7 pagesBasic Concepts On Laboratory Biosafety and BiosecurityKristine PangahinNo ratings yet
- The Immune SystemDocument19 pagesThe Immune Systemmay esteban100% (2)
- ajol-file-journals_82_articles_82639_submission_proof_82639-973-198882-1-10-20121025Document8 pagesajol-file-journals_82_articles_82639_submission_proof_82639-973-198882-1-10-20121025yarach9494No ratings yet
- Food ControlDocument7 pagesFood ControlValentin GhenceaNo ratings yet
- Officejmbfs,+Jmbfs 1626 OyusaglamDocument5 pagesOfficejmbfs,+Jmbfs 1626 OyusaglamWendy SarmientoNo ratings yet
- Food Microbiology 85 (2020) 103282Document8 pagesFood Microbiology 85 (2020) 103282shinchanNo ratings yet
- Example Article IJFMDocument12 pagesExample Article IJFMMarcel HoungbédjiNo ratings yet
- Microbiological assessment of fresh, minimally processed vegetables from open air markets and supermarkets in Luzon, Philippines, for food safetyDocument10 pagesMicrobiological assessment of fresh, minimally processed vegetables from open air markets and supermarkets in Luzon, Philippines, for food safetyiyaNo ratings yet
- 1 Pilihan Hari JugaDocument9 pages1 Pilihan Hari JuganadyasuwayviaNo ratings yet
- Fresh Cut Pineapple As A New Carrier of Probiotic Lactic Acid BacteriaDocument10 pagesFresh Cut Pineapple As A New Carrier of Probiotic Lactic Acid Bacteriahossein alirezaeiNo ratings yet
- Armando Et Al 2011Document10 pagesArmando Et Al 2011Adrian Melgratti JobsonNo ratings yet
- 2018 - Adebayo & Martínez-Carrera - IJ Food Science & TechnologyDocument15 pages2018 - Adebayo & Martínez-Carrera - IJ Food Science & TechnologyAdair Abundis IbarraNo ratings yet
- 90Document11 pages90Eka Nurul HayatiNo ratings yet
- Biological Control: Min Zheng, Jingying Shi, Jian Shi, Qingguo Wang, Yanhua LiDocument7 pagesBiological Control: Min Zheng, Jingying Shi, Jian Shi, Qingguo Wang, Yanhua LiKieu Viet AnhNo ratings yet
- Klaus 2021Document14 pagesKlaus 2021Juan QuitianNo ratings yet
- 7IJEEFUSAUG20197Document8 pages7IJEEFUSAUG20197TJPRC PublicationsNo ratings yet
- C0 Formulir PemohonanDocument5 pagesC0 Formulir PemohonanDiah AyuningrumNo ratings yet
- Escherichia Coli O157:H7, and Listeria Monocytogenes On Fresh LettuceDocument6 pagesEscherichia Coli O157:H7, and Listeria Monocytogenes On Fresh LettuceTanmoy Kumar DeyNo ratings yet
- Antibacterial Mechanism of Lactic Acid On Physiological andDocument6 pagesAntibacterial Mechanism of Lactic Acid On Physiological andlox standardNo ratings yet
- 1 s2.0 S0362028X22084447 MainDocument9 pages1 s2.0 S0362028X22084447 MainTanti 01No ratings yet
- TocoshDocument8 pagesTocoshHansNo ratings yet
- 2021 Antagonistic Activity of Lactic Acid Bacteria Against Phytopathogenic Fungi Isolated From Cherry TomatoDocument10 pages2021 Antagonistic Activity of Lactic Acid Bacteria Against Phytopathogenic Fungi Isolated From Cherry TomatoEduardo Urbano Moraga CáceresNo ratings yet
- Furlaneto-Maia2020 Article AntimicrobialActivityOfEnterocDocument10 pagesFurlaneto-Maia2020 Article AntimicrobialActivityOfEnterocAmanuel AdugnaNo ratings yet
- 45 fr-2021-446 AliDocument9 pages45 fr-2021-446 AliHikmah KimaNo ratings yet
- Isolation of Antibacterial Components From Infusion of Caesalpinia ParaguariensisDocument10 pagesIsolation of Antibacterial Components From Infusion of Caesalpinia ParaguariensisAndres Fernando Silvestre SuarezNo ratings yet
- Nutrients 09 00755Document22 pagesNutrients 09 00755John James MonteverdeNo ratings yet
- Food Microbiology: Meenu N. Perera, Tamar Abuladze, Manrong Li, Joelle Woolston, Alexander SulakvelidzeDocument7 pagesFood Microbiology: Meenu N. Perera, Tamar Abuladze, Manrong Li, Joelle Woolston, Alexander Sulakvelidzeapria siskaNo ratings yet
- The Chemical Preservatives Used To ControlDocument6 pagesThe Chemical Preservatives Used To ControlAmine BenineNo ratings yet
- Food Chemistry: Analytical MethodsDocument5 pagesFood Chemistry: Analytical MethodsFatehatun NoorNo ratings yet
- Reuteri 2-20B and Pediococcus Acidilactici 0-11A Isolated From FuraDocument9 pagesReuteri 2-20B and Pediococcus Acidilactici 0-11A Isolated From FuramostecocNo ratings yet
- 1 s2.0 S0362028X22018543 MainDocument4 pages1 s2.0 S0362028X22018543 MainVNo ratings yet
- Probiotics As Control Agents in AquacultureDocument4 pagesProbiotics As Control Agents in AquacultureBerlin Coldfish BarkerNo ratings yet
- Lactobacillus BrevisDocument21 pagesLactobacillus Brevisshreya singhNo ratings yet
- Pan, X., Chen, F., Wu, T., Tang, H., Dan Zhao, Z. 2009. The Acid, Bile Tolerance and Antimicrobial Property of Lactobacillus Acidophilus NIT. J. Food Control. 20 598-602.Document5 pagesPan, X., Chen, F., Wu, T., Tang, H., Dan Zhao, Z. 2009. The Acid, Bile Tolerance and Antimicrobial Property of Lactobacillus Acidophilus NIT. J. Food Control. 20 598-602.AfdhalRuslanNo ratings yet
- Research Article: in Vitro Evaluation of Probiotic Properties of Lactic Acid BacteriaDocument12 pagesResearch Article: in Vitro Evaluation of Probiotic Properties of Lactic Acid BacteriaNatalija Atanasova-PancevskaNo ratings yet
- Budu BiotekDocument5 pagesBudu BiotekIndra Yusuf PratamaNo ratings yet
- BacteriophagesDocument5 pagesBacteriophagesJames UgbesNo ratings yet
- Characterisation of Bacteriocins Produced by LactoDocument10 pagesCharacterisation of Bacteriocins Produced by Lactokhurry testNo ratings yet
- Franco 2016Document15 pagesFranco 2016RogerioportelaNo ratings yet
- Bacteriocins From Lactic Acid Bacteria Inhibit Food Borne Pathogens.Document7 pagesBacteriocins From Lactic Acid Bacteria Inhibit Food Borne Pathogens.International Organization of Scientific Research (IOSR)No ratings yet
- Invitro Probiotic Evaluation of Lactic Acid BacteriaDocument8 pagesInvitro Probiotic Evaluation of Lactic Acid Bacteria325071067No ratings yet
- EFFECTS OF CALCIUM ASCORBATE AND CALCIUM LACTATE ON QUALITY OF FRESH-CUT PINEAPPLE (Ananas Comosus) (#544974) - 709246Document8 pagesEFFECTS OF CALCIUM ASCORBATE AND CALCIUM LACTATE ON QUALITY OF FRESH-CUT PINEAPPLE (Ananas Comosus) (#544974) - 709246Gaganpreet KaurNo ratings yet
- Journal RosettoDocument9 pagesJournal RosettoBrad PorneaNo ratings yet
- Food Control: Djamel Djenane, Javier Yangüela, Luis Montañés, Mouloud Djerbal, Pedro RoncalésDocument8 pagesFood Control: Djamel Djenane, Javier Yangüela, Luis Montañés, Mouloud Djerbal, Pedro RoncalésEstefaníaNo ratings yet
- Abo-State Et Al., 2015Document7 pagesAbo-State Et Al., 2015Jordana KalineNo ratings yet
- Martínez Et Al 2018Document10 pagesMartínez Et Al 2018Adrian Melgratti JobsonNo ratings yet
- In-Vitro Screening of Antibacterial Activity of Lactic Acid Bacteria Against Common Enteric PathogensDocument6 pagesIn-Vitro Screening of Antibacterial Activity of Lactic Acid Bacteria Against Common Enteric PathogensInternational Medical PublisherNo ratings yet
- Acid LaticoDocument9 pagesAcid LaticoWilson SilvaNo ratings yet
- The Lactic Acid Bacteria (Lab) Activities Against Pathogenic and Spoilage Bacteria On A Dried Salted LoligoDocument8 pagesThe Lactic Acid Bacteria (Lab) Activities Against Pathogenic and Spoilage Bacteria On A Dried Salted LoligocumabuatdownloadNo ratings yet
- In Vitro SourdoughDocument11 pagesIn Vitro SourdoughMaryam JameelahNo ratings yet
- EuglenaDocument8 pagesEuglenaHAROLD HUMBERTO REYES ALVAREZNo ratings yet
- (Allameh, 2012) Isolation, Identification and Characterization of Leuconostoc Mesenteroides As A New Probiotic From Intestine of Snakehead Fish (Channa Striatus) .Document7 pages(Allameh, 2012) Isolation, Identification and Characterization of Leuconostoc Mesenteroides As A New Probiotic From Intestine of Snakehead Fish (Channa Striatus) .José Luis Plaza DoradoNo ratings yet
- Barbosa2013 PDFDocument6 pagesBarbosa2013 PDFSilvia Guerrero GonzalezNo ratings yet
- Holz LAB AntifungalDocument17 pagesHolz LAB AntifungalIstván NagyNo ratings yet
- Ochratoxin A Induced Growth Disturbance, Immune Suppression, and Histopathological Changes of Liver and Kidney of Nil...Document18 pagesOchratoxin A Induced Growth Disturbance, Immune Suppression, and Histopathological Changes of Liver and Kidney of Nil...Abdallah Tag Eldein MansourNo ratings yet
- Safe Practices for Food Processes _ Analysis and Evaluation of Preventive Control Measures for the Control and Reduction_Elimination of Microbial Hazards on Fresh and Fresh-Cut Produce_ List of ReferencesDocument41 pagesSafe Practices for Food Processes _ Analysis and Evaluation of Preventive Control Measures for the Control and Reduction_Elimination of Microbial Hazards on Fresh and Fresh-Cut Produce_ List of ReferencesMohamed ElsawyNo ratings yet
- MicrobiologicalDocument12 pagesMicrobiologicalTrung Truong NhatNo ratings yet
- Preliminary Characterization of BacterioDocument9 pagesPreliminary Characterization of Bacteriomanuomar35No ratings yet
- BacteriocinDocument16 pagesBacteriocintransformers7No ratings yet
- Antioxidant and Antibacterial Activities of Polyphenols From Ethnomedicinal Plants of Burkina FasoDocument6 pagesAntioxidant and Antibacterial Activities of Polyphenols From Ethnomedicinal Plants of Burkina Fasokaori_lawlietNo ratings yet
- Antimicrobial Activities of An Oxygenated Cyclohexanone Derivative Isolated FromDocument36 pagesAntimicrobial Activities of An Oxygenated Cyclohexanone Derivative Isolated FromUtari ZulfianiNo ratings yet
- Nursing Care For A Family Caring For Children With Infectious DiseaseDocument4 pagesNursing Care For A Family Caring For Children With Infectious DiseasekdfhjfhfNo ratings yet
- Endocrine NotesDocument157 pagesEndocrine NotesLaila Cristine CarvalhoNo ratings yet
- Bacteriology Mtap (Part 2)Document10 pagesBacteriology Mtap (Part 2)Joshua CayagoNo ratings yet
- Bacon 2006Document10 pagesBacon 2006Bryan Lloyd LauritoNo ratings yet
- Histology Lab GuideDocument96 pagesHistology Lab GuidevrindaNo ratings yet
- HPV16 Genetic Variation and The Development of Cervical Cancer WorldwideDocument5 pagesHPV16 Genetic Variation and The Development of Cervical Cancer WorldwideQuyết Lê HữuNo ratings yet
- General Biology 2Document5 pagesGeneral Biology 2Kim Ashley Padernal MandronNo ratings yet
- Signaling of Receptor Tyrosine Kinases in The Nucleus: Sally-Anne Stephenson, Inga Mertens-Walker and Adrian HeringtonDocument25 pagesSignaling of Receptor Tyrosine Kinases in The Nucleus: Sally-Anne Stephenson, Inga Mertens-Walker and Adrian HeringtonAyu SafitriNo ratings yet
- Current UIP (Universal Immunization Programme) Schedule in Our StateDocument35 pagesCurrent UIP (Universal Immunization Programme) Schedule in Our StateGopireddy Sivakarthik ReddyNo ratings yet
- Antibiotic ProductionDocument19 pagesAntibiotic ProductionBright OgwoNo ratings yet
- HDN Alias Eritroblastosis FetalisDocument6 pagesHDN Alias Eritroblastosis FetalisOlivia RamadhaniNo ratings yet
- Biological Classification MCQDocument17 pagesBiological Classification MCQNawaraj RokayaNo ratings yet
- Science and Technology Sep June EnglishDocument85 pagesScience and Technology Sep June EnglishJAGADEESH AMNo ratings yet
- CeftolozaneTazobactam A New Option in The TreatmenDocument7 pagesCeftolozaneTazobactam A New Option in The TreatmenAisyah RohieNo ratings yet
- Gene Action and Non-Mendelian InheritanceDocument39 pagesGene Action and Non-Mendelian InheritanceFrank AyrosoNo ratings yet
- Chapter 10: Muscle Tissue: VoluntaryDocument13 pagesChapter 10: Muscle Tissue: VoluntaryClyde BaltazarNo ratings yet
- Effectofberberineon GiardiasisDocument5 pagesEffectofberberineon GiardiasisSummiNo ratings yet
- Quiz RenalDocument81 pagesQuiz RenalMedShare86% (14)
- Avanced Lipoproteins Reserch 2017Document108 pagesAvanced Lipoproteins Reserch 2017Carlos SorianoNo ratings yet
- Arroz Con Mango PDFDocument197 pagesArroz Con Mango PDFVanshika JainNo ratings yet
- HBs Ag Q2Document8 pagesHBs Ag Q2Naveen Kumar MNo ratings yet
- Mutation: Altered Genes: Apago PDF EnhancerDocument5 pagesMutation: Altered Genes: Apago PDF EnhancerAndresNo ratings yet
- The Buzz On Caffeine in Invertebrates Effects On Behavior and Molecular MechanismsDocument9 pagesThe Buzz On Caffeine in Invertebrates Effects On Behavior and Molecular Mechanismsittoiram setagNo ratings yet
- Conceptual Issues in Patenting of Life Forms.Document7 pagesConceptual Issues in Patenting of Life Forms.Jay PatelNo ratings yet
- Antifungal Azole Derivatives and Their Pharmacological Potential: Prospects & RetrospectsDocument14 pagesAntifungal Azole Derivatives and Their Pharmacological Potential: Prospects & RetrospectsRenata YolandaNo ratings yet
- Tetracycline's: Jagir R. Patel Asst Prof Dept. Pharmacology Anand Pharmacy CollegeDocument21 pagesTetracycline's: Jagir R. Patel Asst Prof Dept. Pharmacology Anand Pharmacy CollegeJagirNo ratings yet
- Master Thesis Cell CultureDocument5 pagesMaster Thesis Cell Cultureebonybatesshreveport100% (2)