
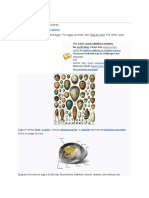
Amniotas vivกparos
Amniotas vivกparos
You might also like
- Chapter 33 - Descriptive EmbryologyDocument12 pagesChapter 33 - Descriptive Embryologyngquvu67% (15)
- Vertebrate Worksheet AnswersDocument5 pagesVertebrate Worksheet AnswersPak RisNo ratings yet
- Animal Kingdom Subphylum Vertebrata Class IXDocument1 pageAnimal Kingdom Subphylum Vertebrata Class IXNandishwara ThonthanalNo ratings yet
- Descriptive Embryology PDFDocument12 pagesDescriptive Embryology PDFMuhammad KhalilNo ratings yet
- Jump To Navigationjump To Search: This Article Is About Biological Eggs. For Eggs As Food, See - For Other Uses, SeeDocument5 pagesJump To Navigationjump To Search: This Article Is About Biological Eggs. For Eggs As Food, See - For Other Uses, SeeewbNo ratings yet
- Animal Kingdom-Subphylum Vertebrata-Class IXDocument2 pagesAnimal Kingdom-Subphylum Vertebrata-Class IXMonika Mehan100% (2)
- Έγγραφο1Document5 pagesΈγγραφο1Pro ProNo ratings yet
- Biology 453 - Comparative Vert. Anatomy WEEK 1, LAB 2: Embryology of Frog & ChickDocument9 pagesBiology 453 - Comparative Vert. Anatomy WEEK 1, LAB 2: Embryology of Frog & ChickPunk Midget-FairyNo ratings yet
- 8-Archaeology and Phylogeny Ascertain Evolution Amniotic Egg-ViviparityDocument10 pages8-Archaeology and Phylogeny Ascertain Evolution Amniotic Egg-ViviparityTeresa Rosales ThamNo ratings yet
- Introduction To FertilizationDocument4 pagesIntroduction To FertilizationSupermarc OlaguirNo ratings yet
- The Eggshell Structure, Composition and MineralizationDocument15 pagesThe Eggshell Structure, Composition and MineralizationydisetiawanNo ratings yet
- SCIENCE YEAR 4 (19.05.2021) Vertebrates and Invertebrates: AnimalsDocument2 pagesSCIENCE YEAR 4 (19.05.2021) Vertebrates and Invertebrates: AnimalsAten KenangaNo ratings yet
- Question Ans On Animal ClassificationDocument3 pagesQuestion Ans On Animal ClassificationIrma HakimNo ratings yet
- Types of Yolk: Structure Specific Origin Fate in Adult Body System FunctionDocument2 pagesTypes of Yolk: Structure Specific Origin Fate in Adult Body System FunctionKobee BacolodNo ratings yet
- Egg As Food Egg (Disambiguation) : Jump To Navigation Jump To SearchDocument7 pagesEgg As Food Egg (Disambiguation) : Jump To Navigation Jump To Searchshreyaz 10No ratings yet
- Early Ontogenesis of The Angelfish, Pterophyllum Scalare Schultze, 1823 (Cichlidae)Document10 pagesEarly Ontogenesis of The Angelfish, Pterophyllum Scalare Schultze, 1823 (Cichlidae)kusuma wardanaNo ratings yet
- Science - ReviewerDocument3 pagesScience - ReviewerTora RaaNo ratings yet
- Birds and Mammals Embryo DevelopmentDocument21 pagesBirds and Mammals Embryo Developmentyui kirigayayuukiNo ratings yet
- Reproduction of EchinodermDocument3 pagesReproduction of EchinodermKesy S. LokaNo ratings yet
- Gen Bio 02 Q4Document7 pagesGen Bio 02 Q4Charles EspiloyNo ratings yet
- 12 - Animal ReproductionDocument32 pages12 - Animal ReproductionMuke MpilwenhleNo ratings yet
- Mind MapDocument2 pagesMind MapBinanana KiutNo ratings yet
- Pudney 1995 PDFDocument39 pagesPudney 1995 PDFKei MlNo ratings yet
- Phylogeny 23Document17 pagesPhylogeny 23Bhavna SharmaNo ratings yet
- Brunet - y - King - 2016 - The Origin of Animal Multicellularity and Cell Differentiation - mmc2Document29 pagesBrunet - y - King - 2016 - The Origin of Animal Multicellularity and Cell Differentiation - mmc2carciclaroNo ratings yet
- Chapter 43 - Animal Reproduction and DevelopmentDocument29 pagesChapter 43 - Animal Reproduction and DevelopmentDóra ImreNo ratings yet
- Regional Specification in The Early Embryo of The Brittle Star Ophiopholis AculeataDocument16 pagesRegional Specification in The Early Embryo of The Brittle Star Ophiopholis AculeataJoElunico QuezadaNo ratings yet
- Lecture - Evolution of AnimalsDocument39 pagesLecture - Evolution of Animalsbenbarb1803No ratings yet
- Asexual Reproduction in InvertebratesDocument11 pagesAsexual Reproduction in Invertebratesaiman maqsood100% (1)
- Burung Matin PDFDocument2 pagesBurung Matin PDFRiskian Alfaro BabtistaNo ratings yet
- Sexual and Asexual Reproduction SeparateDocument2 pagesSexual and Asexual Reproduction SeparateBongani Dean SayedNo ratings yet
- ATELERIX Albiventris ReproductionDocument8 pagesATELERIX Albiventris ReproductionmariaNo ratings yet
- ATELERIX Albiventris Reproduction PDFDocument8 pagesATELERIX Albiventris Reproduction PDFmariaNo ratings yet
- cá sấuDocument57 pagescá sấuHạt Vừng LépNo ratings yet
- Animal EggsDocument14 pagesAnimal EggshosoksnbNo ratings yet
- Normal Embryonic Development in The Clearnose Skate, Raja Eglanteria, With Experimental Observations On Artificial InseminationDocument17 pagesNormal Embryonic Development in The Clearnose Skate, Raja Eglanteria, With Experimental Observations On Artificial InseminationJosé Vélez TacuriNo ratings yet
- Comparative Vertebrate Anatomy (Lab Final Exam) Name: Deanne Marie S. Derecho Date: October 23, 2020 Topic: Reproductive SystemDocument6 pagesComparative Vertebrate Anatomy (Lab Final Exam) Name: Deanne Marie S. Derecho Date: October 23, 2020 Topic: Reproductive SystemShaira CogollodoNo ratings yet
- Biology Labs Online EvolutionDocument4 pagesBiology Labs Online Evolution19claire67No ratings yet
- Parental Care in Fishes (With Diagram) - Vertebrates - Chordata - ZoologyDocument12 pagesParental Care in Fishes (With Diagram) - Vertebrates - Chordata - ZoologyEr Purushottam Pal100% (1)
- 06 GastrulationtxtDocument38 pages06 GastrulationtxtHafidzul HalimNo ratings yet
- Module 4. FertilizationDocument13 pagesModule 4. FertilizationSebastian SmytheNo ratings yet
- PDFDocument13 pagesPDFPranjal ChakrabortyNo ratings yet
- Animal Reproductive ModesDocument33 pagesAnimal Reproductive ModesJessa BelleNo ratings yet
- Chicks Offer Insight Into Origin of FlightDocument16 pagesChicks Offer Insight Into Origin of FlightAlice CalviNo ratings yet
- A Solar SalamanderDocument4 pagesA Solar SalamanderJavier DonosoNo ratings yet
- Insect Reproductive SystemDocument3 pagesInsect Reproductive SystemLiezl Bornales ParreñoNo ratings yet
- 3 EvolutionDocument21 pages3 EvolutionUtkarsh AnandNo ratings yet
- U5 Coral Reproduction BackgroundDocument10 pagesU5 Coral Reproduction BackgroundIsabella Maria Trujillo PulgarNo ratings yet
- VRF 10 001Document7 pagesVRF 10 001Sbastian PratamaNo ratings yet
- Larva: Evolutionary TheoryDocument2 pagesLarva: Evolutionary TheoryEdron Bullo JrNo ratings yet
- Extra Embrional Membrane: Lab Activity VDocument9 pagesExtra Embrional Membrane: Lab Activity VAficko Razaky PratamaNo ratings yet
- Enrichment Activity Gen BioDocument2 pagesEnrichment Activity Gen BioHendrik CondeNo ratings yet
- External and Internal FertilizationDocument2 pagesExternal and Internal FertilizationSISANo ratings yet
- LFSC Paper 1 Last Push WORKBOOK MemoDocument24 pagesLFSC Paper 1 Last Push WORKBOOK MemorebeccakegopotsoeNo ratings yet
- ADocument40 pagesAapi-3826751No ratings yet
- Zoo301lex2 4 Intros Alviar2017Document2 pagesZoo301lex2 4 Intros Alviar2017Mary Rose SantiagoNo ratings yet
- The Eggshell: Structure, Composition and Mineralization: Frontiers in Bioscience January 2012Document16 pagesThe Eggshell: Structure, Composition and Mineralization: Frontiers in Bioscience January 2012John Carlo LayugNo ratings yet
- Reproduction SystemDocument38 pagesReproduction SystemNurfatin AdilaNo ratings yet
- The Eggshell: Structure, Composition and Mineralization: Frontiers in Bioscience January 2012Document16 pagesThe Eggshell: Structure, Composition and Mineralization: Frontiers in Bioscience January 2012Lagasca IrisNo ratings yet
- Pigeons - A Collection of Articles on the Origins, Varieties and Methods of Pigeon KeepingFrom EverandPigeons - A Collection of Articles on the Origins, Varieties and Methods of Pigeon KeepingNo ratings yet
- Petrified Embryology Volume 3: The Frozen Baby Dinosaurs - Spinosaurus dorsojuvencusFrom EverandPetrified Embryology Volume 3: The Frozen Baby Dinosaurs - Spinosaurus dorsojuvencusNo ratings yet
- 424Document107 pages424zulhelmy92No ratings yet
- Bio Zoology 11thDocument247 pagesBio Zoology 11thPradeep ChandarNo ratings yet
- RL 549.3 003 v.4Document337 pagesRL 549.3 003 v.4Hazi TelapokaNo ratings yet
- Zoology TestDocument56 pagesZoology TestHero Khan UsafxaiNo ratings yet
- CK-12 Biology Chapter 17 WorksheetsDocument16 pagesCK-12 Biology Chapter 17 WorksheetsmaleskunNo ratings yet
- Invertebrates Chordata VertebratesDocument62 pagesInvertebrates Chordata VertebratesDao Ming SiNo ratings yet
- Making and Analyzing Cladograms: Name: Nguyen Duc Minh Date: 10/1/2020Document3 pagesMaking and Analyzing Cladograms: Name: Nguyen Duc Minh Date: 10/1/2020Minh Nguyen Duc0% (2)
- Reptile ReproductionDocument5 pagesReptile ReproductionDiyar Ahmad100% (1)
- Zoology 10Th Edition Stephen A Miller 2 All ChapterDocument67 pagesZoology 10Th Edition Stephen A Miller 2 All Chapterdevin.brown498100% (9)
- Reptile Biodiversity STANDARD METHODS FOR INVENTORY AND MONITORING PDFDocument425 pagesReptile Biodiversity STANDARD METHODS FOR INVENTORY AND MONITORING PDFBabi ToroBarros100% (2)
- SeanWilliamsonThesisFinal PDFDocument248 pagesSeanWilliamsonThesisFinal PDFLê Thị Ngọc ĐiệpNo ratings yet
- Introduction To ChordataDocument203 pagesIntroduction To ChordataYhan Brotamonte BoneoNo ratings yet
- (Methods in Molecular Biology 1290) Anoop Kumar, András Simon (Eds.) - Salamanders in Regeneration Research - Methods and ProtocolsDocument346 pages(Methods in Molecular Biology 1290) Anoop Kumar, András Simon (Eds.) - Salamanders in Regeneration Research - Methods and ProtocolsDiana ToroNo ratings yet
- Vertebrate Life 10Th Edition Edition Pough Ebook Full ChapterDocument51 pagesVertebrate Life 10Th Edition Edition Pough Ebook Full Chapterpeter.rivera411100% (6)
- The History of Animal EvolutionDocument11 pagesThe History of Animal EvolutionMong GulongNo ratings yet
- Class Amphibia MammaliaDocument7 pagesClass Amphibia MammaliaKristella DraheimNo ratings yet
- REPTILESDocument52 pagesREPTILESBerces BelleNo ratings yet
- Kamus AntropologiDocument89 pagesKamus Antropologipandupandawa100% (2)
- Animal EvolutionDocument6 pagesAnimal EvolutionYsabella PolanaNo ratings yet
- Class ReptiliaDocument119 pagesClass ReptiliaArwa ZakriaNo ratings yet
- Reptiles I PDFDocument16 pagesReptiles I PDFNitgma DcNo ratings yet
- Zelfstudie Van Blastula Naar 3 KiemlagenDocument18 pagesZelfstudie Van Blastula Naar 3 KiemlagenCheyenne ProvoostNo ratings yet
- Powerpoint Lectures For: Biology, Seventh EditionDocument124 pagesPowerpoint Lectures For: Biology, Seventh Editionxo_simpledream100% (1)
- 4 - Tetrapods IDocument20 pages4 - Tetrapods Ibhavikrao7605No ratings yet
- ReptilesDocument2 pagesReptilesapi-417823294No ratings yet
- Tetrapods + AmniotesDocument40 pagesTetrapods + Amnioteskfagg0tNo ratings yet
- Fossils in Utah Booklet 1Document21 pagesFossils in Utah Booklet 1api-218354277No ratings yet
- 105 Lab 7 Paleozoic F21.ONLINE ONLYDocument10 pages105 Lab 7 Paleozoic F21.ONLINE ONLYJimmy DoyleNo ratings yet
- GrandmotherfishDocument18 pagesGrandmotherfishe.mirikolNo ratings yet




























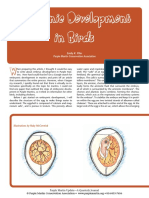




























































Download as pdf or txt
You might also like
- Chapter 33 - Descriptive EmbryologyDocument12 pagesChapter 33 - Descriptive Embryologyngquvu67% (15)
- Vertebrate Worksheet AnswersDocument5 pagesVertebrate Worksheet AnswersPak RisNo ratings yet
- Animal Kingdom Subphylum Vertebrata Class IXDocument1 pageAnimal Kingdom Subphylum Vertebrata Class IXNandishwara ThonthanalNo ratings yet
- Descriptive Embryology PDFDocument12 pagesDescriptive Embryology PDFMuhammad KhalilNo ratings yet
- Jump To Navigationjump To Search: This Article Is About Biological Eggs. For Eggs As Food, See - For Other Uses, SeeDocument5 pagesJump To Navigationjump To Search: This Article Is About Biological Eggs. For Eggs As Food, See - For Other Uses, SeeewbNo ratings yet
- Animal Kingdom-Subphylum Vertebrata-Class IXDocument2 pagesAnimal Kingdom-Subphylum Vertebrata-Class IXMonika Mehan100% (2)
- Έγγραφο1Document5 pagesΈγγραφο1Pro ProNo ratings yet
- Biology 453 - Comparative Vert. Anatomy WEEK 1, LAB 2: Embryology of Frog & ChickDocument9 pagesBiology 453 - Comparative Vert. Anatomy WEEK 1, LAB 2: Embryology of Frog & ChickPunk Midget-FairyNo ratings yet
- 8-Archaeology and Phylogeny Ascertain Evolution Amniotic Egg-ViviparityDocument10 pages8-Archaeology and Phylogeny Ascertain Evolution Amniotic Egg-ViviparityTeresa Rosales ThamNo ratings yet
- Introduction To FertilizationDocument4 pagesIntroduction To FertilizationSupermarc OlaguirNo ratings yet
- The Eggshell Structure, Composition and MineralizationDocument15 pagesThe Eggshell Structure, Composition and MineralizationydisetiawanNo ratings yet
- SCIENCE YEAR 4 (19.05.2021) Vertebrates and Invertebrates: AnimalsDocument2 pagesSCIENCE YEAR 4 (19.05.2021) Vertebrates and Invertebrates: AnimalsAten KenangaNo ratings yet
- Question Ans On Animal ClassificationDocument3 pagesQuestion Ans On Animal ClassificationIrma HakimNo ratings yet
- Types of Yolk: Structure Specific Origin Fate in Adult Body System FunctionDocument2 pagesTypes of Yolk: Structure Specific Origin Fate in Adult Body System FunctionKobee BacolodNo ratings yet
- Egg As Food Egg (Disambiguation) : Jump To Navigation Jump To SearchDocument7 pagesEgg As Food Egg (Disambiguation) : Jump To Navigation Jump To Searchshreyaz 10No ratings yet
- Early Ontogenesis of The Angelfish, Pterophyllum Scalare Schultze, 1823 (Cichlidae)Document10 pagesEarly Ontogenesis of The Angelfish, Pterophyllum Scalare Schultze, 1823 (Cichlidae)kusuma wardanaNo ratings yet
- Science - ReviewerDocument3 pagesScience - ReviewerTora RaaNo ratings yet
- Birds and Mammals Embryo DevelopmentDocument21 pagesBirds and Mammals Embryo Developmentyui kirigayayuukiNo ratings yet
- Reproduction of EchinodermDocument3 pagesReproduction of EchinodermKesy S. LokaNo ratings yet
- Gen Bio 02 Q4Document7 pagesGen Bio 02 Q4Charles EspiloyNo ratings yet
- 12 - Animal ReproductionDocument32 pages12 - Animal ReproductionMuke MpilwenhleNo ratings yet
- Mind MapDocument2 pagesMind MapBinanana KiutNo ratings yet
- Pudney 1995 PDFDocument39 pagesPudney 1995 PDFKei MlNo ratings yet
- Phylogeny 23Document17 pagesPhylogeny 23Bhavna SharmaNo ratings yet
- Brunet - y - King - 2016 - The Origin of Animal Multicellularity and Cell Differentiation - mmc2Document29 pagesBrunet - y - King - 2016 - The Origin of Animal Multicellularity and Cell Differentiation - mmc2carciclaroNo ratings yet
- Chapter 43 - Animal Reproduction and DevelopmentDocument29 pagesChapter 43 - Animal Reproduction and DevelopmentDóra ImreNo ratings yet
- Regional Specification in The Early Embryo of The Brittle Star Ophiopholis AculeataDocument16 pagesRegional Specification in The Early Embryo of The Brittle Star Ophiopholis AculeataJoElunico QuezadaNo ratings yet
- Lecture - Evolution of AnimalsDocument39 pagesLecture - Evolution of Animalsbenbarb1803No ratings yet
- Asexual Reproduction in InvertebratesDocument11 pagesAsexual Reproduction in Invertebratesaiman maqsood100% (1)
- Burung Matin PDFDocument2 pagesBurung Matin PDFRiskian Alfaro BabtistaNo ratings yet
- Sexual and Asexual Reproduction SeparateDocument2 pagesSexual and Asexual Reproduction SeparateBongani Dean SayedNo ratings yet
- ATELERIX Albiventris ReproductionDocument8 pagesATELERIX Albiventris ReproductionmariaNo ratings yet
- ATELERIX Albiventris Reproduction PDFDocument8 pagesATELERIX Albiventris Reproduction PDFmariaNo ratings yet
- cá sấuDocument57 pagescá sấuHạt Vừng LépNo ratings yet
- Animal EggsDocument14 pagesAnimal EggshosoksnbNo ratings yet
- Normal Embryonic Development in The Clearnose Skate, Raja Eglanteria, With Experimental Observations On Artificial InseminationDocument17 pagesNormal Embryonic Development in The Clearnose Skate, Raja Eglanteria, With Experimental Observations On Artificial InseminationJosé Vélez TacuriNo ratings yet
- Comparative Vertebrate Anatomy (Lab Final Exam) Name: Deanne Marie S. Derecho Date: October 23, 2020 Topic: Reproductive SystemDocument6 pagesComparative Vertebrate Anatomy (Lab Final Exam) Name: Deanne Marie S. Derecho Date: October 23, 2020 Topic: Reproductive SystemShaira CogollodoNo ratings yet
- Biology Labs Online EvolutionDocument4 pagesBiology Labs Online Evolution19claire67No ratings yet
- Parental Care in Fishes (With Diagram) - Vertebrates - Chordata - ZoologyDocument12 pagesParental Care in Fishes (With Diagram) - Vertebrates - Chordata - ZoologyEr Purushottam Pal100% (1)
- 06 GastrulationtxtDocument38 pages06 GastrulationtxtHafidzul HalimNo ratings yet
- Module 4. FertilizationDocument13 pagesModule 4. FertilizationSebastian SmytheNo ratings yet
- PDFDocument13 pagesPDFPranjal ChakrabortyNo ratings yet
- Animal Reproductive ModesDocument33 pagesAnimal Reproductive ModesJessa BelleNo ratings yet
- Chicks Offer Insight Into Origin of FlightDocument16 pagesChicks Offer Insight Into Origin of FlightAlice CalviNo ratings yet
- A Solar SalamanderDocument4 pagesA Solar SalamanderJavier DonosoNo ratings yet
- Insect Reproductive SystemDocument3 pagesInsect Reproductive SystemLiezl Bornales ParreñoNo ratings yet
- 3 EvolutionDocument21 pages3 EvolutionUtkarsh AnandNo ratings yet
- U5 Coral Reproduction BackgroundDocument10 pagesU5 Coral Reproduction BackgroundIsabella Maria Trujillo PulgarNo ratings yet
- VRF 10 001Document7 pagesVRF 10 001Sbastian PratamaNo ratings yet
- Larva: Evolutionary TheoryDocument2 pagesLarva: Evolutionary TheoryEdron Bullo JrNo ratings yet
- Extra Embrional Membrane: Lab Activity VDocument9 pagesExtra Embrional Membrane: Lab Activity VAficko Razaky PratamaNo ratings yet
- Enrichment Activity Gen BioDocument2 pagesEnrichment Activity Gen BioHendrik CondeNo ratings yet
- External and Internal FertilizationDocument2 pagesExternal and Internal FertilizationSISANo ratings yet
- LFSC Paper 1 Last Push WORKBOOK MemoDocument24 pagesLFSC Paper 1 Last Push WORKBOOK MemorebeccakegopotsoeNo ratings yet
- ADocument40 pagesAapi-3826751No ratings yet
- Zoo301lex2 4 Intros Alviar2017Document2 pagesZoo301lex2 4 Intros Alviar2017Mary Rose SantiagoNo ratings yet
- The Eggshell: Structure, Composition and Mineralization: Frontiers in Bioscience January 2012Document16 pagesThe Eggshell: Structure, Composition and Mineralization: Frontiers in Bioscience January 2012John Carlo LayugNo ratings yet
- Reproduction SystemDocument38 pagesReproduction SystemNurfatin AdilaNo ratings yet
- The Eggshell: Structure, Composition and Mineralization: Frontiers in Bioscience January 2012Document16 pagesThe Eggshell: Structure, Composition and Mineralization: Frontiers in Bioscience January 2012Lagasca IrisNo ratings yet
- Pigeons - A Collection of Articles on the Origins, Varieties and Methods of Pigeon KeepingFrom EverandPigeons - A Collection of Articles on the Origins, Varieties and Methods of Pigeon KeepingNo ratings yet
- Petrified Embryology Volume 3: The Frozen Baby Dinosaurs - Spinosaurus dorsojuvencusFrom EverandPetrified Embryology Volume 3: The Frozen Baby Dinosaurs - Spinosaurus dorsojuvencusNo ratings yet
- 424Document107 pages424zulhelmy92No ratings yet
- Bio Zoology 11thDocument247 pagesBio Zoology 11thPradeep ChandarNo ratings yet
- RL 549.3 003 v.4Document337 pagesRL 549.3 003 v.4Hazi TelapokaNo ratings yet
- Zoology TestDocument56 pagesZoology TestHero Khan UsafxaiNo ratings yet
- CK-12 Biology Chapter 17 WorksheetsDocument16 pagesCK-12 Biology Chapter 17 WorksheetsmaleskunNo ratings yet
- Invertebrates Chordata VertebratesDocument62 pagesInvertebrates Chordata VertebratesDao Ming SiNo ratings yet
- Making and Analyzing Cladograms: Name: Nguyen Duc Minh Date: 10/1/2020Document3 pagesMaking and Analyzing Cladograms: Name: Nguyen Duc Minh Date: 10/1/2020Minh Nguyen Duc0% (2)
- Reptile ReproductionDocument5 pagesReptile ReproductionDiyar Ahmad100% (1)
- Zoology 10Th Edition Stephen A Miller 2 All ChapterDocument67 pagesZoology 10Th Edition Stephen A Miller 2 All Chapterdevin.brown498100% (9)
- Reptile Biodiversity STANDARD METHODS FOR INVENTORY AND MONITORING PDFDocument425 pagesReptile Biodiversity STANDARD METHODS FOR INVENTORY AND MONITORING PDFBabi ToroBarros100% (2)
- SeanWilliamsonThesisFinal PDFDocument248 pagesSeanWilliamsonThesisFinal PDFLê Thị Ngọc ĐiệpNo ratings yet
- Introduction To ChordataDocument203 pagesIntroduction To ChordataYhan Brotamonte BoneoNo ratings yet
- (Methods in Molecular Biology 1290) Anoop Kumar, András Simon (Eds.) - Salamanders in Regeneration Research - Methods and ProtocolsDocument346 pages(Methods in Molecular Biology 1290) Anoop Kumar, András Simon (Eds.) - Salamanders in Regeneration Research - Methods and ProtocolsDiana ToroNo ratings yet
- Vertebrate Life 10Th Edition Edition Pough Ebook Full ChapterDocument51 pagesVertebrate Life 10Th Edition Edition Pough Ebook Full Chapterpeter.rivera411100% (6)
- The History of Animal EvolutionDocument11 pagesThe History of Animal EvolutionMong GulongNo ratings yet
- Class Amphibia MammaliaDocument7 pagesClass Amphibia MammaliaKristella DraheimNo ratings yet
- REPTILESDocument52 pagesREPTILESBerces BelleNo ratings yet
- Kamus AntropologiDocument89 pagesKamus Antropologipandupandawa100% (2)
- Animal EvolutionDocument6 pagesAnimal EvolutionYsabella PolanaNo ratings yet
- Class ReptiliaDocument119 pagesClass ReptiliaArwa ZakriaNo ratings yet
- Reptiles I PDFDocument16 pagesReptiles I PDFNitgma DcNo ratings yet
- Zelfstudie Van Blastula Naar 3 KiemlagenDocument18 pagesZelfstudie Van Blastula Naar 3 KiemlagenCheyenne ProvoostNo ratings yet
- Powerpoint Lectures For: Biology, Seventh EditionDocument124 pagesPowerpoint Lectures For: Biology, Seventh Editionxo_simpledream100% (1)
- 4 - Tetrapods IDocument20 pages4 - Tetrapods Ibhavikrao7605No ratings yet
- ReptilesDocument2 pagesReptilesapi-417823294No ratings yet
- Tetrapods + AmniotesDocument40 pagesTetrapods + Amnioteskfagg0tNo ratings yet
- Fossils in Utah Booklet 1Document21 pagesFossils in Utah Booklet 1api-218354277No ratings yet
- 105 Lab 7 Paleozoic F21.ONLINE ONLYDocument10 pages105 Lab 7 Paleozoic F21.ONLINE ONLYJimmy DoyleNo ratings yet
- GrandmotherfishDocument18 pagesGrandmotherfishe.mirikolNo ratings yet