Download as pdf or txt
You might also like
- Sherris Medical Microbiology Seventh Edition Ebook PDF Version 7thDocument61 pagesSherris Medical Microbiology Seventh Edition Ebook PDF Version 7ththeresa.whitley258100% (45)
- CHAPTER 12 Lymphatic SystemDocument139 pagesCHAPTER 12 Lymphatic SystemMaria Christilaine ReyesNo ratings yet
- Clasificarea Bacteriilor I.Gram Pozitivi 1.aerobi A.CociDocument7 pagesClasificarea Bacteriilor I.Gram Pozitivi 1.aerobi A.Cocialtereu100% (6)
- Lecture 13 Tumor ImmunologyDocument99 pagesLecture 13 Tumor Immunologyمحمود محمد100% (1)
- Correlation Between Antimicrobial Resistance, Biofilm Formation, and Virulence Determinants in Uropathogenic Escherichia Coli From Egyptian HospitalDocument22 pagesCorrelation Between Antimicrobial Resistance, Biofilm Formation, and Virulence Determinants in Uropathogenic Escherichia Coli From Egyptian Hospitald4rkgr455No ratings yet
- I - Inhibition of Streptococcus Mutans Biofilms With Bacterial-Derived Outer Membrane VesiclesDocument12 pagesI - Inhibition of Streptococcus Mutans Biofilms With Bacterial-Derived Outer Membrane VesiclesHime IbaraNo ratings yet
- Potential Properties of Lactobacillus Plantarum F-10 As A Bio-Control Strategy For Wound InfectionsDocument14 pagesPotential Properties of Lactobacillus Plantarum F-10 As A Bio-Control Strategy For Wound InfectionsIgor BaltaNo ratings yet
- Current Promising Opportunities of Treating Ploymicro DiseaesDocument29 pagesCurrent Promising Opportunities of Treating Ploymicro Diseaesczikkely23No ratings yet
- Immunoinformatic For Potential Vaccine DiscoveryDocument23 pagesImmunoinformatic For Potential Vaccine DiscoveryDesye MeleseNo ratings yet
- Modulating The Transcriptomic Profile of Multidrug-Resistant Klebsiella Pneumoniae Biofilm Formation by Antibiotics in Combination With Zinc SulfateDocument12 pagesModulating The Transcriptomic Profile of Multidrug-Resistant Klebsiella Pneumoniae Biofilm Formation by Antibiotics in Combination With Zinc Sulfated4rkgr455No ratings yet
- Dual-Function Antimicrobial-Antibiofilm Peptide Hybrid To Tackle Biofilm-Forming Staphylococcus EpidermidisDocument19 pagesDual-Function Antimicrobial-Antibiofilm Peptide Hybrid To Tackle Biofilm-Forming Staphylococcus Epidermidisd4rkgr455No ratings yet
- J of Applied Microbiology - 2016 - Ferrer - Effect of Antibiotics On Biofilm Inhibition and Induction Measured by Real TimeDocument11 pagesJ of Applied Microbiology - 2016 - Ferrer - Effect of Antibiotics On Biofilm Inhibition and Induction Measured by Real TimeRin ChanNo ratings yet
- Determining Effects of Nitrate, Arginine, and Ferrous On Antibiotic Recalcitrance of Clinical Strains of Pseudomonas Aeruginosa in Biofilm-Inspired Alginate EncapsulatesDocument11 pagesDetermining Effects of Nitrate, Arginine, and Ferrous On Antibiotic Recalcitrance of Clinical Strains of Pseudomonas Aeruginosa in Biofilm-Inspired Alginate Encapsulatesd4rkgr455No ratings yet
- AntibioticsDocument12 pagesAntibioticsValentinaNo ratings yet
- 2 PMID 37992081 BiofilmDocument15 pages2 PMID 37992081 Biofilmajaya basnetNo ratings yet
- Lecture 1Document17 pagesLecture 1mheni4979No ratings yet
- Bio Film: Sowmya Subramanian, Ryan C. Huiszoon, Sangwook Chu, William E. Bentley, Reza GhodssiDocument16 pagesBio Film: Sowmya Subramanian, Ryan C. Huiszoon, Sangwook Chu, William E. Bentley, Reza GhodssiOussama El BouadiNo ratings yet
- Jurnal 14Document23 pagesJurnal 14evefilerespulNo ratings yet
- s12917 022 03501 3Document11 pagess12917 022 03501 3huangfang921001No ratings yet
- Review Molecular Interactions in BiofilmsDocument13 pagesReview Molecular Interactions in Biofilmsnaima aminaNo ratings yet
- Kumar 2017Document9 pagesKumar 2017farah moallaNo ratings yet
- An Optimized Anti-Adherence and Anti-Biofilm Assay Case Study of Zinc Oxide Nanoparticles Versus MRSA BiofilmDocument6 pagesAn Optimized Anti-Adherence and Anti-Biofilm Assay Case Study of Zinc Oxide Nanoparticles Versus MRSA BiofilmKareem IsmailNo ratings yet
- Novel Bacteriophage Specific Against Staphylococcus Epidermidis and With Antibiofilm ActivityDocument11 pagesNovel Bacteriophage Specific Against Staphylococcus Epidermidis and With Antibiofilm ActivityZaenab AzzahraNo ratings yet
- Understanding Antimicrobial Susceptibility Profile of Finegoldia Magna An Insight To An Untrodden PathDocument5 pagesUnderstanding Antimicrobial Susceptibility Profile of Finegoldia Magna An Insight To An Untrodden Pathd4rkgr455No ratings yet
- Abiofilm: A Resource of Anti-Biofilm Agents and Their Potential Implications in Targeting Antibiotic Drug ResistanceDocument7 pagesAbiofilm: A Resource of Anti-Biofilm Agents and Their Potential Implications in Targeting Antibiotic Drug ResistanceBeshoy RafatNo ratings yet
- Link Between MIC and Antimicrobial TherapyDocument8 pagesLink Between MIC and Antimicrobial TherapySahar WaqarNo ratings yet
- Plants 11 01429 v3Document20 pagesPlants 11 01429 v3Parmeshver BhagatNo ratings yet
- Current Anti-Biofilm Strategies and Potential of Antioxidants in Biofilm ControlDocument10 pagesCurrent Anti-Biofilm Strategies and Potential of Antioxidants in Biofilm ControlBeshoy RafatNo ratings yet
- Effect of Anti-Biofilm Potential of Different Medicinal Plants: ReviewDocument9 pagesEffect of Anti-Biofilm Potential of Different Medicinal Plants: ReviewmoupriyanagNo ratings yet
- Biofilm Intracelular 2020Document9 pagesBiofilm Intracelular 2020Gabriela ChaconNo ratings yet
- Antimicrobial and Antibiofilm Potential of Injectable Platelet RichDocument6 pagesAntimicrobial and Antibiofilm Potential of Injectable Platelet RichNurul Fajriah YunitaNo ratings yet
- Bacterial Biofilm Its Composition Formation and Role in Human Infections1Document15 pagesBacterial Biofilm Its Composition Formation and Role in Human Infections1Zaky RaihanNo ratings yet
- The Role of Bacterial Biofilms in Device-Associated InfectionDocument10 pagesThe Role of Bacterial Biofilms in Device-Associated InfectionLorena Bellan DomicianoNo ratings yet
- Review of Biofilm Inhibiting PhytochemicalsDocument26 pagesReview of Biofilm Inhibiting PhytochemicalsMichal SladekNo ratings yet
- JMB 31 1 1Document7 pagesJMB 31 1 1icrgNo ratings yet
- Aplicación de Campos MagnéticosDocument12 pagesAplicación de Campos MagnéticosGustavoNo ratings yet
- Engineering Bacteria For Cancer Immunotherapy - ScienceDirectDocument16 pagesEngineering Bacteria For Cancer Immunotherapy - ScienceDirectlauracarrio2007No ratings yet
- Combating Multidrug Resistance: The Potential of Antimicrobial Peptides and Biofilm ChallengesDocument29 pagesCombating Multidrug Resistance: The Potential of Antimicrobial Peptides and Biofilm ChallengesInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Isolation of Pathogens Causing Sepsis Pus and Infected Wounds From Critical Care Unit A Retrospective Study - 2Document8 pagesIsolation of Pathogens Causing Sepsis Pus and Infected Wounds From Critical Care Unit A Retrospective Study - 2Omoding EmmanuelNo ratings yet
- IJBMS Volume 23 Issue 5 Pages 691-698Document8 pagesIJBMS Volume 23 Issue 5 Pages 691-698amanda.thaisNo ratings yet
- Microbial Biofilms 2020Document6 pagesMicrobial Biofilms 2020AlexandraTacoNo ratings yet
- BBCT2Document15 pagesBBCT2Kajelcha FikaduNo ratings yet
- Artigo NutriçãoDocument4 pagesArtigo Nutriçãoannakah18No ratings yet
- Microbiota Intestinal e Inmunidad. Review.2020Document15 pagesMicrobiota Intestinal e Inmunidad. Review.2020hacek357No ratings yet
- Pharmaceuticals 16 00741Document17 pagesPharmaceuticals 16 00741Mayara BernickerNo ratings yet
- Genetic Determinants of Macrolide and Tetracycline Resistance in Penicillin Non-Susceptible Streptococcus Pneumoniae Isolates From People Living With HIV in Dar Es Salaam, TanzaniaDocument7 pagesGenetic Determinants of Macrolide and Tetracycline Resistance in Penicillin Non-Susceptible Streptococcus Pneumoniae Isolates From People Living With HIV in Dar Es Salaam, Tanzaniad4rkgr455No ratings yet
- Staphylococcus BiofilmsDocument20 pagesStaphylococcus BiofilmsOssian89No ratings yet
- Detection of Antibiotic Resistance and Bio Film-Producing Ability of Staphylococcus Species in Clinical IsolatesDocument5 pagesDetection of Antibiotic Resistance and Bio Film-Producing Ability of Staphylococcus Species in Clinical Isolatesafnan jumaaNo ratings yet
- Biofilm Formation by The Interaction of Fungi With Varoius BacteriaDocument10 pagesBiofilm Formation by The Interaction of Fungi With Varoius BacteriaLaborat MenurNo ratings yet
- ZeamaysDocument9 pagesZeamaysGurinder SinghNo ratings yet
- Lab Report Antimicrobial Susceptibility Testing MethodsDocument19 pagesLab Report Antimicrobial Susceptibility Testing MethodsAlyaa Athira100% (1)
- Microbiota Manipulation As An Emerging Concept in Cancer TherapyDocument9 pagesMicrobiota Manipulation As An Emerging Concept in Cancer TherapypherretyNo ratings yet
- 2023 Article 2897Document13 pages2023 Article 2897Robert StryjakNo ratings yet
- 2Document12 pages2Annu OjhaNo ratings yet
- Antimicrobial Resistance To Disinfectants in Biofilms: P.Araújo, M.Lemos, F.Mergulhão, L. Melo and M.SimõesDocument9 pagesAntimicrobial Resistance To Disinfectants in Biofilms: P.Araújo, M.Lemos, F.Mergulhão, L. Melo and M.SimõesNatalija Atanasova-PancevskaNo ratings yet
- Microbial Ecology of The Skin in The Era of Metagenomics and Molecular MicrobiologyDocument35 pagesMicrobial Ecology of The Skin in The Era of Metagenomics and Molecular MicrobiologyIsabelaEstradaRodriguezNo ratings yet
- Microbiota Ocular e Inflamación IntraocularDocument12 pagesMicrobiota Ocular e Inflamación IntraocularDiego QuinteroNo ratings yet
- 2021 Article 624Document18 pages2021 Article 624icrgNo ratings yet
- Jurnal 5Document7 pagesJurnal 5kalstarNo ratings yet
- Plants: A Path Forward: Promoting Microbial-Based Methods in The Control of Invasive Plant SpeciesDocument13 pagesPlants: A Path Forward: Promoting Microbial-Based Methods in The Control of Invasive Plant SpeciesChristina MathewNo ratings yet
- Final Synopsis Sandeep PDFDocument10 pagesFinal Synopsis Sandeep PDFKalyani GNo ratings yet
- Annurev Anchem 091522 103827Document21 pagesAnnurev Anchem 091522 103827Robert StryjakNo ratings yet
- Pseudomonas Aeruginosa - Formarea BiofilmuluiDocument25 pagesPseudomonas Aeruginosa - Formarea BiofilmuluiRadu ZubaciNo ratings yet
- Nature's Answer to Viral Threats: Understanding the Potency of Plant-Based AntiviralsFrom EverandNature's Answer to Viral Threats: Understanding the Potency of Plant-Based AntiviralsNo ratings yet
- Improvement in Neisseria Gonorrhoeae Culture Rates by Bedside Inoculation and Incubation at A Clinic For Sexually Transmitted InfectionsDocument9 pagesImprovement in Neisseria Gonorrhoeae Culture Rates by Bedside Inoculation and Incubation at A Clinic For Sexually Transmitted Infectionsd4rkgr455No ratings yet
- Impact of Influenza Virus Infection On Lung Microbiome in Adults With Severe PneumoniaDocument12 pagesImpact of Influenza Virus Infection On Lung Microbiome in Adults With Severe Pneumoniad4rkgr455No ratings yet
- External Ventriculostomy-Associated Infection Reduction After Updating A Care BundleDocument10 pagesExternal Ventriculostomy-Associated Infection Reduction After Updating A Care Bundled4rkgr455No ratings yet
- An Autopsy Case of Infective Aortic Aneurysm With Pasteurella Multocida Infection Clinicopathological Appearance and A Review of LiteraturesDocument7 pagesAn Autopsy Case of Infective Aortic Aneurysm With Pasteurella Multocida Infection Clinicopathological Appearance and A Review of Literaturesd4rkgr455No ratings yet
- Next-Generation Sequencing in The Diagnosis of Neurobrucellosis A Case Series of Eight Consecutive PatientsDocument8 pagesNext-Generation Sequencing in The Diagnosis of Neurobrucellosis A Case Series of Eight Consecutive Patientsd4rkgr455No ratings yet
- Risk Factors Associated With Colistin Resistance in Carbapenemase-Producing Enterobacterales A Multicenter Study From A Low-Income CountryDocument9 pagesRisk Factors Associated With Colistin Resistance in Carbapenemase-Producing Enterobacterales A Multicenter Study From A Low-Income Countryd4rkgr455No ratings yet
- Genomic Epidemiology Reveals Multiple Mechanisms of Linezolid Resistance in Clinical Enterococci in ChinaDocument11 pagesGenomic Epidemiology Reveals Multiple Mechanisms of Linezolid Resistance in Clinical Enterococci in Chinad4rkgr455No ratings yet
- Multidrug-Resistant Gram-Negative Bacterial Infections and Associated Factors in A Kenyan Intensive Care Unit A Cross-Sectional StudyDocument11 pagesMultidrug-Resistant Gram-Negative Bacterial Infections and Associated Factors in A Kenyan Intensive Care Unit A Cross-Sectional Studyd4rkgr455No ratings yet
- Tetracyclines Resistance in Mycoplasma and Ureaplasma Urogenital Isolates Derived From Human A Systematic Review and Meta-AnalysisDocument12 pagesTetracyclines Resistance in Mycoplasma and Ureaplasma Urogenital Isolates Derived From Human A Systematic Review and Meta-Analysisd4rkgr455No ratings yet
- Modulating The Transcriptomic Profile of Multidrug-Resistant Klebsiella Pneumoniae Biofilm Formation by Antibiotics in Combination With Zinc SulfateDocument12 pagesModulating The Transcriptomic Profile of Multidrug-Resistant Klebsiella Pneumoniae Biofilm Formation by Antibiotics in Combination With Zinc Sulfated4rkgr455No ratings yet
- Dual-Function Antimicrobial-Antibiofilm Peptide Hybrid To Tackle Biofilm-Forming Staphylococcus EpidermidisDocument19 pagesDual-Function Antimicrobial-Antibiofilm Peptide Hybrid To Tackle Biofilm-Forming Staphylococcus Epidermidisd4rkgr455No ratings yet
- Genomic Characterization of Colistin-Resistant Klebsiella Pneumoniae Isolated From Intensive Care Unit Patients in EgyptDocument16 pagesGenomic Characterization of Colistin-Resistant Klebsiella Pneumoniae Isolated From Intensive Care Unit Patients in Egyptd4rkgr455No ratings yet
- Bigger Problems From Smaller Colonies Emergence of Antibiotic-Tolerant Small Colony Variants of Mycobacterium Avium Complex in MAC-pulmonary Disease PatientsDocument16 pagesBigger Problems From Smaller Colonies Emergence of Antibiotic-Tolerant Small Colony Variants of Mycobacterium Avium Complex in MAC-pulmonary Disease Patientsd4rkgr455No ratings yet
- Obsd Faq49Document318 pagesObsd Faq49d4rkgr455No ratings yet
- Phenotypic and Genotypic Analysis of Antimicrobial Resistance and Population Structure of Gastroenteritis-Related Aeromonas IsolatesDocument8 pagesPhenotypic and Genotypic Analysis of Antimicrobial Resistance and Population Structure of Gastroenteritis-Related Aeromonas Isolatesd4rkgr455No ratings yet
- Differences in Clinical Outcomes of Bloodstream Infections Caused by Klebsiella Aerogenes, Klebsiella Pneumoniae and Enterobacter Cloacae A Multicentre Cohort StudyDocument11 pagesDifferences in Clinical Outcomes of Bloodstream Infections Caused by Klebsiella Aerogenes, Klebsiella Pneumoniae and Enterobacter Cloacae A Multicentre Cohort Studyd4rkgr455No ratings yet
- Conversion RPM G CentrifugaDocument1 pageConversion RPM G CentrifugaEsaú E RodriguezNo ratings yet
- Coronavirus (COVID-19) Testing - Statistics and Research - Our World in DataDocument89 pagesCoronavirus (COVID-19) Testing - Statistics and Research - Our World in Datahah266611No ratings yet
- Covid-19 RT PCR Test Sample Type: Method UsedDocument1 pageCovid-19 RT PCR Test Sample Type: Method UsedAditya UrkudeNo ratings yet
- Eul 0515 202 00 Covid19 Coronavirus Real Time PCR Kit IfuDocument28 pagesEul 0515 202 00 Covid19 Coronavirus Real Time PCR Kit IfuashishvaidNo ratings yet
- Cells Activity QuestionsDocument2 pagesCells Activity QuestionsMusab AlbarbariNo ratings yet
- Anaerobic Blood Culture Bottle IFU en July 18, 2012Document3 pagesAnaerobic Blood Culture Bottle IFU en July 18, 2012Hadi AlbitarNo ratings yet
- COVID-19 Update For Nurses: Greetings From The QA Team!!Document8 pagesCOVID-19 Update For Nurses: Greetings From The QA Team!!Mable CNo ratings yet
- Interpretation of Hepatitis SerologyDocument2 pagesInterpretation of Hepatitis SerologyTugasbu CicikNo ratings yet
- Answers To Genetics Practice Exam 1Document15 pagesAnswers To Genetics Practice Exam 1cowboyNo ratings yet
- COVID-19 Leaky Vaccine Hesitancy v2.5Document26 pagesCOVID-19 Leaky Vaccine Hesitancy v2.5XaceboppNo ratings yet
- Mass Gatherings Evidence ReviewDocument72 pagesMass Gatherings Evidence ReviewRogueNo ratings yet
- COVID-19 Pandemic in The PhilippinesDocument89 pagesCOVID-19 Pandemic in The PhilippinesAnthony BanateNo ratings yet
- Allegranzi-Benedetta HH Escmid PDFDocument92 pagesAllegranzi-Benedetta HH Escmid PDFNishanth.sNo ratings yet
- Covid 19Document22 pagesCovid 19imad khanNo ratings yet
- ASHP COVID 19 Evidence TableDocument138 pagesASHP COVID 19 Evidence TableOrchid Limoso LozanoNo ratings yet
- Laboratory Test Report: Test Name Result Sars-Cov-2 (RDRP Gene)Document1 pageLaboratory Test Report: Test Name Result Sars-Cov-2 (RDRP Gene)srinathNo ratings yet
- MeningitisDocument12 pagesMeningitisDany100% (5)
- Orchid Virus Diseases in Taiwan and Their Control StrategiesDocument54 pagesOrchid Virus Diseases in Taiwan and Their Control StrategiesPham Lanphuong100% (1)
- 19-12-2020 HMB EnglishDocument12 pages19-12-2020 HMB EnglishPushNo ratings yet
- Immun e System: Chapter 31.2 Page 885Document21 pagesImmun e System: Chapter 31.2 Page 885JanaNo ratings yet
- BMS533 Lab PracticalDocument3 pagesBMS533 Lab PracticalYUSNURDALILA DALINo ratings yet
- Sars-Cov-2 (Covid 19) Detection (Qualitative) by Real Time RT PCRDocument3 pagesSars-Cov-2 (Covid 19) Detection (Qualitative) by Real Time RT PCRMabelann VazNo ratings yet
- Hind III Haemophilus InfluenzaeDocument2 pagesHind III Haemophilus InfluenzaeJazmin Nicole AbanesNo ratings yet
- Guidance On The Management of HIV-infected Healthcare Workers in The Philippines 2017Document21 pagesGuidance On The Management of HIV-infected Healthcare Workers in The Philippines 2017psmid-webNo ratings yet
- NewEarth - University COVID-19 Intelligence BriefDocument47 pagesNewEarth - University COVID-19 Intelligence BriefTony Lambert100% (1)
- Inspekcija Krompir, NAKDocument38 pagesInspekcija Krompir, NAKgandra80No ratings yet
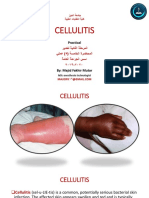
- Cellulitis: PracticalDocument11 pagesCellulitis: PracticalAli dhyaaNo ratings yet