
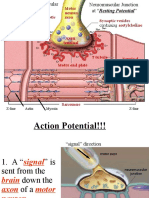
(K Leak Channels) Establish The Resting Potential of A Neuron
(K Leak Channels) Establish The Resting Potential of A Neuron
You might also like
- Intro To FSM Handouts BWDocument30 pagesIntro To FSM Handouts BWroberto_celio_2100% (5)
- Resting Membrane, Graded, Action Potentials AtfDocument4 pagesResting Membrane, Graded, Action Potentials AtfdaphneNo ratings yet
- 9 Draw StructDocument6 pages9 Draw StructkalloliNo ratings yet
- PharmacologyDocument271 pagesPharmacologysweta suman100% (4)
- A History of Membrane Transport and Bioenergetics PDFDocument383 pagesA History of Membrane Transport and Bioenergetics PDFEdmilson RodriguesNo ratings yet
- V. A. Shepherd (Auth.), Alexander G. Volkov (Eds.) - Plant Electrophysiology - Methods and Cell Electrophysiology-Springer-Verlag Berlin Heidelberg (2012) PDFDocument375 pagesV. A. Shepherd (Auth.), Alexander G. Volkov (Eds.) - Plant Electrophysiology - Methods and Cell Electrophysiology-Springer-Verlag Berlin Heidelberg (2012) PDFMaria Elsa RodriguezNo ratings yet
- Potential - Best Compatibility ModeDocument14 pagesPotential - Best Compatibility ModeDivya TripathyNo ratings yet
- Nervous SystemDocument10 pagesNervous SystemkasheeNo ratings yet
- UntitledDocument86 pagesUntitledMichael Saganya LuhagaNo ratings yet
- Nervous System FundamentalsDocument29 pagesNervous System FundamentalsSarosh MajeedNo ratings yet
- Potencial de AcciónDocument23 pagesPotencial de AcciónEstefania Olmos MoralesNo ratings yet
- Biolistrik: Irfiansyah IrwadiDocument39 pagesBiolistrik: Irfiansyah IrwadiRakhmatul Binti SulistyaNo ratings yet
- Pharmacology - Anti-Arrhythmias 2019 PDFDocument11 pagesPharmacology - Anti-Arrhythmias 2019 PDFpurletpunkNo ratings yet
- Biology 946: Unit 9 - Control & Regulation Section 1 - Nervous SystemDocument47 pagesBiology 946: Unit 9 - Control & Regulation Section 1 - Nervous SystemverysedatedxNo ratings yet
- 01the Nerve Impulsepc-2Document42 pages01the Nerve Impulsepc-2Fwesa evelyn MukimbaNo ratings yet
- Synapse: Is The Point of Contact Between A Neuron andDocument31 pagesSynapse: Is The Point of Contact Between A Neuron andCharlie Mejos MercedNo ratings yet
- Neuronal Transmission (Prof. Ieva)Document38 pagesNeuronal Transmission (Prof. Ieva)Anna Rahmania SariNo ratings yet
- 10.3 The Nerve ImpulseDocument20 pages10.3 The Nerve ImpulseHaruka SuzukiNo ratings yet
- Ionic Channel Na - K Pump Na VGC K VGC: Mambrane Potential With Value (MV) Inside ChargeDocument8 pagesIonic Channel Na - K Pump Na VGC K VGC: Mambrane Potential With Value (MV) Inside ChargekasheeNo ratings yet
- Study Notes For Biology Exam 2Document19 pagesStudy Notes For Biology Exam 2YaffaAliNo ratings yet
- NS 1 Electrical SignalingDocument29 pagesNS 1 Electrical Signalingyamanuel25No ratings yet
- Comparison Between Neuronal Action Potential and Action Potentials in Other CellsDocument26 pagesComparison Between Neuronal Action Potential and Action Potentials in Other CellsmaryamNo ratings yet
- Neurons and The Physiology of The Nervous SystemDocument34 pagesNeurons and The Physiology of The Nervous SystemEmanNo ratings yet
- Neuron Communication LectureDocument31 pagesNeuron Communication LectureJennifer Isom Schmidtke, Ph.D.100% (1)
- Introduction To Neuroscience-Communication Within NeuronsDocument34 pagesIntroduction To Neuroscience-Communication Within NeuronsGonzalo BustosNo ratings yet
- Nerve ImpulseDocument7 pagesNerve ImpulseyadnispamNo ratings yet
- E Valency Ion Ion: A - Ve Value Means A Net Flow Into The CellDocument5 pagesE Valency Ion Ion: A - Ve Value Means A Net Flow Into The Cellravi2likeNo ratings yet
- 4. Transport Across Cell Membranes - CopyDocument35 pages4. Transport Across Cell Membranes - Copyowotiajoshua7No ratings yet
- Human Anatomy and PhysiologyDocument42 pagesHuman Anatomy and PhysiologySantino AwetNo ratings yet
- Biopsych Slide 4Document40 pagesBiopsych Slide 4JImmy LauNo ratings yet
- TA Session 6 2Document38 pagesTA Session 6 2viviannaNo ratings yet
- Nervous System: AP BiologyDocument30 pagesNervous System: AP Biologyjulie rainesNo ratings yet
- Genesis of Resting Membrane PotentialDocument9 pagesGenesis of Resting Membrane PotentialSyauqat AthirNo ratings yet
- Stimulates Ca ") .: 1997 Pharmacology Semester 1 Page 1 of 6Document6 pagesStimulates Ca ") .: 1997 Pharmacology Semester 1 Page 1 of 6ravi2likeNo ratings yet
- 4 Tejidos Excitables 2019Document70 pages4 Tejidos Excitables 2019Martita JuárezNo ratings yet
- Y1S1 Membrane PotentialsDocument57 pagesY1S1 Membrane PotentialsdwataNo ratings yet
- 21.1 Action PotentialDocument43 pages21.1 Action PotentialNazirah Arba'inNo ratings yet
- Z-Line Z-Line Actin Myosin: Na+ Na+ Na+ Na+ Na+ Na+Document35 pagesZ-Line Z-Line Actin Myosin: Na+ Na+ Na+ Na+ Na+ Na+dvenumohanNo ratings yet
- PEX-03-01-Pot de Repaus PDFDocument8 pagesPEX-03-01-Pot de Repaus PDFAlexandru BurdeniucNo ratings yet
- L 7 Action PotentialDocument27 pagesL 7 Action PotentialsNo ratings yet
- Bioelectric Potential: Dr. Anju Jha Mbbs. Md. PGDMCHDocument63 pagesBioelectric Potential: Dr. Anju Jha Mbbs. Md. PGDMCHzohaibNo ratings yet
- Exercice 3 Actividad 1Document8 pagesExercice 3 Actividad 1EVELYN BRIGGITE AVILES SANMARTINNo ratings yet
- Pex 03 01Document8 pagesPex 03 01Jose Maria RojasNo ratings yet
- MD-2 Neuron'sDocument36 pagesMD-2 Neuron'svincenzoNo ratings yet
- L6 ActionPotential Basis SLIDESDocument4 pagesL6 ActionPotential Basis SLIDESxicoblendxxwyshNo ratings yet
- Electrophysiology Cardiovascular 2010Document77 pagesElectrophysiology Cardiovascular 2010Palupi Setyo HapsariNo ratings yet
- 0 NS 5 Nerve ImpulsesDocument16 pages0 NS 5 Nerve ImpulsesbadamsmithNo ratings yet
- Action Potential FinalDocument28 pagesAction Potential FinalKawish AliNo ratings yet
- BBMS3011 Synaptic Dysfunction and DepressionDocument29 pagesBBMS3011 Synaptic Dysfunction and Depressionkeven319hk4304No ratings yet
- 2021 Lecture 3 Cardiac Physiology NewDocument41 pages2021 Lecture 3 Cardiac Physiology NewMajd HusseinNo ratings yet
- Neural Signaling: The Membrane Potential: Lesson 9Document30 pagesNeural Signaling: The Membrane Potential: Lesson 9saavanakumarNo ratings yet
- Physiology 2 Membrane PotentialDocument18 pagesPhysiology 2 Membrane Potentialdrkhan58No ratings yet
- Gusgus 1Document8 pagesGusgus 1Mateo MoncayoNo ratings yet
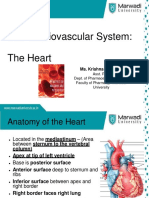
- The Cardiovascular System: The Heart: Part B: Prepared by Barbara Heard, Atlantic Cape Community CollegeDocument80 pagesThe Cardiovascular System: The Heart: Part B: Prepared by Barbara Heard, Atlantic Cape Community CollegeBrianna PinchinatNo ratings yet
- Action PotentialDocument74 pagesAction PotentialAnbreena RasoolNo ratings yet
- B5W4 Renal Regulation of Potassium Homeostasis STUDENTDocument29 pagesB5W4 Renal Regulation of Potassium Homeostasis STUDENTInhoNo ratings yet
- Action Potentials and Synapses HandoutsDocument6 pagesAction Potentials and Synapses HandoutsKelly TrainorNo ratings yet
- 5-Generation and Propagation of Action Potential (2016-17) (Amended)Document21 pages5-Generation and Propagation of Action Potential (2016-17) (Amended)Wai Kwong ChiuNo ratings yet
- Membrane Potential & Action Potential - SZ (Final Term)Document34 pagesMembrane Potential & Action Potential - SZ (Final Term)Tanvir FahimNo ratings yet
- BME 301: Biomedical Sensors: Lecture Note 3: Bioelectric Potentials and Biopotential ElectrodesDocument67 pagesBME 301: Biomedical Sensors: Lecture Note 3: Bioelectric Potentials and Biopotential ElectrodesDewi Cahya FitriNo ratings yet
- 01Document8 pages01Dany BerlinNo ratings yet
- 1B Nazvaira Al Marwah Nenden 1084 EX3 ACT1Document8 pages1B Nazvaira Al Marwah Nenden 1084 EX3 ACT1naspaheheNo ratings yet
- PhysioEx Exercise 3 Activity 1 - Subkelompok 4Document8 pagesPhysioEx Exercise 3 Activity 1 - Subkelompok 4Syarifina PutriNo ratings yet
- In the Wake of the Jomon: Stone Age Mariners and a Voyage Across the PacificFrom EverandIn the Wake of the Jomon: Stone Age Mariners and a Voyage Across the PacificRating: 5 out of 5 stars5/5 (2)
- pectoral chorsiaDocument30 pagespectoral chorsiakalloliNo ratings yet
- jogyallapur revenue mapDocument1 pagejogyallapur revenue mapkalloliNo ratings yet
- protDocument2 pagesprotkalloliNo ratings yet
- 5 PrinciplesDocument2 pages5 PrincipleskalloliNo ratings yet
- tadasinakoppa revenueDocument1 pagetadasinakoppa revenuekalloliNo ratings yet
- 9 EliminationDocument5 pages9 EliminationkalloliNo ratings yet
- 8 MetabolismDocument3 pages8 MetabolismkalloliNo ratings yet
- 1 Enzymes Why They Are Efficient and SpecificDocument7 pages1 Enzymes Why They Are Efficient and SpecifickalloliNo ratings yet
- Pages from Physiology Integrated Approach-3Document1 pagePages from Physiology Integrated Approach-3kalloliNo ratings yet
- 3 Sigma and PaiDocument1 page3 Sigma and PaikalloliNo ratings yet
- 2 Kinetics of Catlyzed ReactionsDocument4 pages2 Kinetics of Catlyzed ReactionskalloliNo ratings yet
- Ashirvad uPVC Casing PipesDocument2 pagesAshirvad uPVC Casing PipeskalloliNo ratings yet
- 6 Plasma CurvesDocument3 pages6 Plasma CurveskalloliNo ratings yet
- Transfer of Building Works To PWOs and PSUsDocument1 pageTransfer of Building Works To PWOs and PSUskalloliNo ratings yet
- Pages From David R. Klein - Organic Chemistry As A Second Language - First Semester Topics-Wiley (2017)Document1 pagePages From David R. Klein - Organic Chemistry As A Second Language - First Semester Topics-Wiley (2017)kalloliNo ratings yet
- Hyperconjugation: The Reactions of Alkenes - The Stereochemistry of Addition ReactionsDocument2 pagesHyperconjugation: The Reactions of Alkenes - The Stereochemistry of Addition ReactionskalloliNo ratings yet
- 3 Ezyme InhibitionDocument3 pages3 Ezyme InhibitionkalloliNo ratings yet
- 2007 Imso ShortanswerDocument10 pages2007 Imso ShortanswerkalloliNo ratings yet
- DPP-1 Circle Ques @motion LecturesDocument1 pageDPP-1 Circle Ques @motion LectureskalloliNo ratings yet
- Pages From TB - 9th EdDocument1 pagePages From TB - 9th EdkalloliNo ratings yet
- DPP-2 Complex-Number Ques @motion LecturesDocument1 pageDPP-2 Complex-Number Ques @motion LectureskalloliNo ratings yet
- DPP-4 Complex-Number Ques @motion LecturesDocument1 pageDPP-4 Complex-Number Ques @motion LectureskalloliNo ratings yet
- 12.3 Structural Representations of Organic Compounds 12.3.1 Complete, Condensed and Bond-Line Structural FormulasDocument3 pages12.3 Structural Representations of Organic Compounds 12.3.1 Complete, Condensed and Bond-Line Structural FormulaskalloliNo ratings yet
- Problem 4.11: Which Would You Expect To Be More Stable: (CH) C or (CF) C ? Why?Document2 pagesProblem 4.11: Which Would You Expect To Be More Stable: (CH) C or (CF) C ? Why?kalloliNo ratings yet
- Func Tional Groups: Heteroatoms-Atoms Other Than Carbon or Hydrogen. Common Heteroatoms Are NitroDocument3 pagesFunc Tional Groups: Heteroatoms-Atoms Other Than Carbon or Hydrogen. Common Heteroatoms Are NitrokalloliNo ratings yet
- Rigidity of Double /triple Bonds: C H H H H H C H H H H C H H H CDocument7 pagesRigidity of Double /triple Bonds: C H H H H H C H H H H C H H H CkalloliNo ratings yet
- The Bonds in Ethene: P P P S SP SP P SPDocument1 pageThe Bonds in Ethene: P P P S SP SP P SPkalloliNo ratings yet
- The Bonds in Ethyne: SP P P P SP S P PDocument1 pageThe Bonds in Ethyne: SP P P P SP S P PkalloliNo ratings yet
- Reso 3Document2 pagesReso 3kalloliNo ratings yet
- Chapter 10 Endocrine SystemDocument8 pagesChapter 10 Endocrine SystemCriscia Lene OlatNo ratings yet
- Cell Communication 1Document10 pagesCell Communication 1api-277471896No ratings yet
- Drugs Used in Disorders of Endocrine System Ppt. Book (Lectures 1-6)Document467 pagesDrugs Used in Disorders of Endocrine System Ppt. Book (Lectures 1-6)Marc Imhotep Cray, M.D.100% (1)
- PharmacologyDocument51 pagesPharmacologyAndy MirandaNo ratings yet
- Revised M SC Zool NEP23!1!2Document108 pagesRevised M SC Zool NEP23!1!2shuklasd3221No ratings yet
- 7 Revesion PDFDocument80 pages7 Revesion PDFMd Sakil AminNo ratings yet
- Mechanisms of Hormone Action NotesDocument3 pagesMechanisms of Hormone Action Notesapi-390361165No ratings yet
- Module 1 Intro PcolDocument21 pagesModule 1 Intro PcolCygme Neile TacusalmeNo ratings yet
- TB cellsignalfrq 65696c9ef1baf2.65696ca0f11c56.93457589 加水印Document11 pagesTB cellsignalfrq 65696c9ef1baf2.65696ca0f11c56.93457589 加水印y77811766No ratings yet
- Cyclic GMP Synthesis, Metabolism, and FunctionDocument353 pagesCyclic GMP Synthesis, Metabolism, and FunctionpawnammalNo ratings yet
- Homo ToxicologyDocument279 pagesHomo ToxicologyRafael MontavaNo ratings yet
- AI Pharma 2 (Edited MCQS)Document11 pagesAI Pharma 2 (Edited MCQS)raneemalnaamnehNo ratings yet
- Biomolecules and Cells NotesDocument81 pagesBiomolecules and Cells NotesLily RobinsonNo ratings yet
- Biochem II Mid Sem 2 MCQs Answered SamiDocument339 pagesBiochem II Mid Sem 2 MCQs Answered Samicarolinegatwiri08No ratings yet
- BIOC1020 Course Document - Outline - SSCDocument4 pagesBIOC1020 Course Document - Outline - SSCbrownjudeaaNo ratings yet
- PharmaDocument147 pagesPharmaDrChauhanNo ratings yet
- Second Messenger SystemsDocument4 pagesSecond Messenger SystemsNatalia Evtikhova EvtikhovNo ratings yet
- Cell SignalingDocument43 pagesCell SignalingRIFAM zNo ratings yet
- Preliminary Term Examination Cytogenetics 001Document16 pagesPreliminary Term Examination Cytogenetics 001MICHAEL JOHN AGUILARNo ratings yet
- Receptor Lecture 1Document24 pagesReceptor Lecture 1Bachelor Gaming CommunityNo ratings yet
- 02 Hormones PDFDocument8 pages02 Hormones PDFKaram SaadNo ratings yet
- BIOM1050: Receptors, Neurons, Synapses & Nervous Systems Cell Communication & Receptor FamiliesDocument12 pagesBIOM1050: Receptors, Neurons, Synapses & Nervous Systems Cell Communication & Receptor FamiliesmichelleNo ratings yet
- Komunikasi Sel PDFDocument64 pagesKomunikasi Sel PDFMuhammad RizaldiNo ratings yet
- Cell SignalingDocument87 pagesCell SignalingZaw Linn PhyoNo ratings yet
- Cell SignalingDocument38 pagesCell SignalingTukai KulkarniNo ratings yet
- Modeling Cellular Signaling Systems: An Abstraction-Refinement ApproachDocument5 pagesModeling Cellular Signaling Systems: An Abstraction-Refinement ApproachdhermitNo ratings yet





















































































































Download as pdf or txt
You might also like
- Intro To FSM Handouts BWDocument30 pagesIntro To FSM Handouts BWroberto_celio_2100% (5)
- Resting Membrane, Graded, Action Potentials AtfDocument4 pagesResting Membrane, Graded, Action Potentials AtfdaphneNo ratings yet
- 9 Draw StructDocument6 pages9 Draw StructkalloliNo ratings yet
- PharmacologyDocument271 pagesPharmacologysweta suman100% (4)
- A History of Membrane Transport and Bioenergetics PDFDocument383 pagesA History of Membrane Transport and Bioenergetics PDFEdmilson RodriguesNo ratings yet
- V. A. Shepherd (Auth.), Alexander G. Volkov (Eds.) - Plant Electrophysiology - Methods and Cell Electrophysiology-Springer-Verlag Berlin Heidelberg (2012) PDFDocument375 pagesV. A. Shepherd (Auth.), Alexander G. Volkov (Eds.) - Plant Electrophysiology - Methods and Cell Electrophysiology-Springer-Verlag Berlin Heidelberg (2012) PDFMaria Elsa RodriguezNo ratings yet
- Potential - Best Compatibility ModeDocument14 pagesPotential - Best Compatibility ModeDivya TripathyNo ratings yet
- Nervous SystemDocument10 pagesNervous SystemkasheeNo ratings yet
- UntitledDocument86 pagesUntitledMichael Saganya LuhagaNo ratings yet
- Nervous System FundamentalsDocument29 pagesNervous System FundamentalsSarosh MajeedNo ratings yet
- Potencial de AcciónDocument23 pagesPotencial de AcciónEstefania Olmos MoralesNo ratings yet
- Biolistrik: Irfiansyah IrwadiDocument39 pagesBiolistrik: Irfiansyah IrwadiRakhmatul Binti SulistyaNo ratings yet
- Pharmacology - Anti-Arrhythmias 2019 PDFDocument11 pagesPharmacology - Anti-Arrhythmias 2019 PDFpurletpunkNo ratings yet
- Biology 946: Unit 9 - Control & Regulation Section 1 - Nervous SystemDocument47 pagesBiology 946: Unit 9 - Control & Regulation Section 1 - Nervous SystemverysedatedxNo ratings yet
- 01the Nerve Impulsepc-2Document42 pages01the Nerve Impulsepc-2Fwesa evelyn MukimbaNo ratings yet
- Synapse: Is The Point of Contact Between A Neuron andDocument31 pagesSynapse: Is The Point of Contact Between A Neuron andCharlie Mejos MercedNo ratings yet
- Neuronal Transmission (Prof. Ieva)Document38 pagesNeuronal Transmission (Prof. Ieva)Anna Rahmania SariNo ratings yet
- 10.3 The Nerve ImpulseDocument20 pages10.3 The Nerve ImpulseHaruka SuzukiNo ratings yet
- Ionic Channel Na - K Pump Na VGC K VGC: Mambrane Potential With Value (MV) Inside ChargeDocument8 pagesIonic Channel Na - K Pump Na VGC K VGC: Mambrane Potential With Value (MV) Inside ChargekasheeNo ratings yet
- Study Notes For Biology Exam 2Document19 pagesStudy Notes For Biology Exam 2YaffaAliNo ratings yet
- NS 1 Electrical SignalingDocument29 pagesNS 1 Electrical Signalingyamanuel25No ratings yet
- Comparison Between Neuronal Action Potential and Action Potentials in Other CellsDocument26 pagesComparison Between Neuronal Action Potential and Action Potentials in Other CellsmaryamNo ratings yet
- Neurons and The Physiology of The Nervous SystemDocument34 pagesNeurons and The Physiology of The Nervous SystemEmanNo ratings yet
- Neuron Communication LectureDocument31 pagesNeuron Communication LectureJennifer Isom Schmidtke, Ph.D.100% (1)
- Introduction To Neuroscience-Communication Within NeuronsDocument34 pagesIntroduction To Neuroscience-Communication Within NeuronsGonzalo BustosNo ratings yet
- Nerve ImpulseDocument7 pagesNerve ImpulseyadnispamNo ratings yet
- E Valency Ion Ion: A - Ve Value Means A Net Flow Into The CellDocument5 pagesE Valency Ion Ion: A - Ve Value Means A Net Flow Into The Cellravi2likeNo ratings yet
- 4. Transport Across Cell Membranes - CopyDocument35 pages4. Transport Across Cell Membranes - Copyowotiajoshua7No ratings yet
- Human Anatomy and PhysiologyDocument42 pagesHuman Anatomy and PhysiologySantino AwetNo ratings yet
- Biopsych Slide 4Document40 pagesBiopsych Slide 4JImmy LauNo ratings yet
- TA Session 6 2Document38 pagesTA Session 6 2viviannaNo ratings yet
- Nervous System: AP BiologyDocument30 pagesNervous System: AP Biologyjulie rainesNo ratings yet
- Genesis of Resting Membrane PotentialDocument9 pagesGenesis of Resting Membrane PotentialSyauqat AthirNo ratings yet
- Stimulates Ca ") .: 1997 Pharmacology Semester 1 Page 1 of 6Document6 pagesStimulates Ca ") .: 1997 Pharmacology Semester 1 Page 1 of 6ravi2likeNo ratings yet
- 4 Tejidos Excitables 2019Document70 pages4 Tejidos Excitables 2019Martita JuárezNo ratings yet
- Y1S1 Membrane PotentialsDocument57 pagesY1S1 Membrane PotentialsdwataNo ratings yet
- 21.1 Action PotentialDocument43 pages21.1 Action PotentialNazirah Arba'inNo ratings yet
- Z-Line Z-Line Actin Myosin: Na+ Na+ Na+ Na+ Na+ Na+Document35 pagesZ-Line Z-Line Actin Myosin: Na+ Na+ Na+ Na+ Na+ Na+dvenumohanNo ratings yet
- PEX-03-01-Pot de Repaus PDFDocument8 pagesPEX-03-01-Pot de Repaus PDFAlexandru BurdeniucNo ratings yet
- L 7 Action PotentialDocument27 pagesL 7 Action PotentialsNo ratings yet
- Bioelectric Potential: Dr. Anju Jha Mbbs. Md. PGDMCHDocument63 pagesBioelectric Potential: Dr. Anju Jha Mbbs. Md. PGDMCHzohaibNo ratings yet
- Exercice 3 Actividad 1Document8 pagesExercice 3 Actividad 1EVELYN BRIGGITE AVILES SANMARTINNo ratings yet
- Pex 03 01Document8 pagesPex 03 01Jose Maria RojasNo ratings yet
- MD-2 Neuron'sDocument36 pagesMD-2 Neuron'svincenzoNo ratings yet
- L6 ActionPotential Basis SLIDESDocument4 pagesL6 ActionPotential Basis SLIDESxicoblendxxwyshNo ratings yet
- Electrophysiology Cardiovascular 2010Document77 pagesElectrophysiology Cardiovascular 2010Palupi Setyo HapsariNo ratings yet
- 0 NS 5 Nerve ImpulsesDocument16 pages0 NS 5 Nerve ImpulsesbadamsmithNo ratings yet
- Action Potential FinalDocument28 pagesAction Potential FinalKawish AliNo ratings yet
- BBMS3011 Synaptic Dysfunction and DepressionDocument29 pagesBBMS3011 Synaptic Dysfunction and Depressionkeven319hk4304No ratings yet
- 2021 Lecture 3 Cardiac Physiology NewDocument41 pages2021 Lecture 3 Cardiac Physiology NewMajd HusseinNo ratings yet
- Neural Signaling: The Membrane Potential: Lesson 9Document30 pagesNeural Signaling: The Membrane Potential: Lesson 9saavanakumarNo ratings yet
- Physiology 2 Membrane PotentialDocument18 pagesPhysiology 2 Membrane Potentialdrkhan58No ratings yet
- Gusgus 1Document8 pagesGusgus 1Mateo MoncayoNo ratings yet
- The Cardiovascular System: The Heart: Part B: Prepared by Barbara Heard, Atlantic Cape Community CollegeDocument80 pagesThe Cardiovascular System: The Heart: Part B: Prepared by Barbara Heard, Atlantic Cape Community CollegeBrianna PinchinatNo ratings yet
- Action PotentialDocument74 pagesAction PotentialAnbreena RasoolNo ratings yet
- B5W4 Renal Regulation of Potassium Homeostasis STUDENTDocument29 pagesB5W4 Renal Regulation of Potassium Homeostasis STUDENTInhoNo ratings yet
- Action Potentials and Synapses HandoutsDocument6 pagesAction Potentials and Synapses HandoutsKelly TrainorNo ratings yet
- 5-Generation and Propagation of Action Potential (2016-17) (Amended)Document21 pages5-Generation and Propagation of Action Potential (2016-17) (Amended)Wai Kwong ChiuNo ratings yet
- Membrane Potential & Action Potential - SZ (Final Term)Document34 pagesMembrane Potential & Action Potential - SZ (Final Term)Tanvir FahimNo ratings yet
- BME 301: Biomedical Sensors: Lecture Note 3: Bioelectric Potentials and Biopotential ElectrodesDocument67 pagesBME 301: Biomedical Sensors: Lecture Note 3: Bioelectric Potentials and Biopotential ElectrodesDewi Cahya FitriNo ratings yet
- 01Document8 pages01Dany BerlinNo ratings yet
- 1B Nazvaira Al Marwah Nenden 1084 EX3 ACT1Document8 pages1B Nazvaira Al Marwah Nenden 1084 EX3 ACT1naspaheheNo ratings yet
- PhysioEx Exercise 3 Activity 1 - Subkelompok 4Document8 pagesPhysioEx Exercise 3 Activity 1 - Subkelompok 4Syarifina PutriNo ratings yet
- In the Wake of the Jomon: Stone Age Mariners and a Voyage Across the PacificFrom EverandIn the Wake of the Jomon: Stone Age Mariners and a Voyage Across the PacificRating: 5 out of 5 stars5/5 (2)
- pectoral chorsiaDocument30 pagespectoral chorsiakalloliNo ratings yet
- jogyallapur revenue mapDocument1 pagejogyallapur revenue mapkalloliNo ratings yet
- protDocument2 pagesprotkalloliNo ratings yet
- 5 PrinciplesDocument2 pages5 PrincipleskalloliNo ratings yet
- tadasinakoppa revenueDocument1 pagetadasinakoppa revenuekalloliNo ratings yet
- 9 EliminationDocument5 pages9 EliminationkalloliNo ratings yet
- 8 MetabolismDocument3 pages8 MetabolismkalloliNo ratings yet
- 1 Enzymes Why They Are Efficient and SpecificDocument7 pages1 Enzymes Why They Are Efficient and SpecifickalloliNo ratings yet
- Pages from Physiology Integrated Approach-3Document1 pagePages from Physiology Integrated Approach-3kalloliNo ratings yet
- 3 Sigma and PaiDocument1 page3 Sigma and PaikalloliNo ratings yet
- 2 Kinetics of Catlyzed ReactionsDocument4 pages2 Kinetics of Catlyzed ReactionskalloliNo ratings yet
- Ashirvad uPVC Casing PipesDocument2 pagesAshirvad uPVC Casing PipeskalloliNo ratings yet
- 6 Plasma CurvesDocument3 pages6 Plasma CurveskalloliNo ratings yet
- Transfer of Building Works To PWOs and PSUsDocument1 pageTransfer of Building Works To PWOs and PSUskalloliNo ratings yet
- Pages From David R. Klein - Organic Chemistry As A Second Language - First Semester Topics-Wiley (2017)Document1 pagePages From David R. Klein - Organic Chemistry As A Second Language - First Semester Topics-Wiley (2017)kalloliNo ratings yet
- Hyperconjugation: The Reactions of Alkenes - The Stereochemistry of Addition ReactionsDocument2 pagesHyperconjugation: The Reactions of Alkenes - The Stereochemistry of Addition ReactionskalloliNo ratings yet
- 3 Ezyme InhibitionDocument3 pages3 Ezyme InhibitionkalloliNo ratings yet
- 2007 Imso ShortanswerDocument10 pages2007 Imso ShortanswerkalloliNo ratings yet
- DPP-1 Circle Ques @motion LecturesDocument1 pageDPP-1 Circle Ques @motion LectureskalloliNo ratings yet
- Pages From TB - 9th EdDocument1 pagePages From TB - 9th EdkalloliNo ratings yet
- DPP-2 Complex-Number Ques @motion LecturesDocument1 pageDPP-2 Complex-Number Ques @motion LectureskalloliNo ratings yet
- DPP-4 Complex-Number Ques @motion LecturesDocument1 pageDPP-4 Complex-Number Ques @motion LectureskalloliNo ratings yet
- 12.3 Structural Representations of Organic Compounds 12.3.1 Complete, Condensed and Bond-Line Structural FormulasDocument3 pages12.3 Structural Representations of Organic Compounds 12.3.1 Complete, Condensed and Bond-Line Structural FormulaskalloliNo ratings yet
- Problem 4.11: Which Would You Expect To Be More Stable: (CH) C or (CF) C ? Why?Document2 pagesProblem 4.11: Which Would You Expect To Be More Stable: (CH) C or (CF) C ? Why?kalloliNo ratings yet
- Func Tional Groups: Heteroatoms-Atoms Other Than Carbon or Hydrogen. Common Heteroatoms Are NitroDocument3 pagesFunc Tional Groups: Heteroatoms-Atoms Other Than Carbon or Hydrogen. Common Heteroatoms Are NitrokalloliNo ratings yet
- Rigidity of Double /triple Bonds: C H H H H H C H H H H C H H H CDocument7 pagesRigidity of Double /triple Bonds: C H H H H H C H H H H C H H H CkalloliNo ratings yet
- The Bonds in Ethene: P P P S SP SP P SPDocument1 pageThe Bonds in Ethene: P P P S SP SP P SPkalloliNo ratings yet
- The Bonds in Ethyne: SP P P P SP S P PDocument1 pageThe Bonds in Ethyne: SP P P P SP S P PkalloliNo ratings yet
- Reso 3Document2 pagesReso 3kalloliNo ratings yet
- Chapter 10 Endocrine SystemDocument8 pagesChapter 10 Endocrine SystemCriscia Lene OlatNo ratings yet
- Cell Communication 1Document10 pagesCell Communication 1api-277471896No ratings yet
- Drugs Used in Disorders of Endocrine System Ppt. Book (Lectures 1-6)Document467 pagesDrugs Used in Disorders of Endocrine System Ppt. Book (Lectures 1-6)Marc Imhotep Cray, M.D.100% (1)
- PharmacologyDocument51 pagesPharmacologyAndy MirandaNo ratings yet
- Revised M SC Zool NEP23!1!2Document108 pagesRevised M SC Zool NEP23!1!2shuklasd3221No ratings yet
- 7 Revesion PDFDocument80 pages7 Revesion PDFMd Sakil AminNo ratings yet
- Mechanisms of Hormone Action NotesDocument3 pagesMechanisms of Hormone Action Notesapi-390361165No ratings yet
- Module 1 Intro PcolDocument21 pagesModule 1 Intro PcolCygme Neile TacusalmeNo ratings yet
- TB cellsignalfrq 65696c9ef1baf2.65696ca0f11c56.93457589 加水印Document11 pagesTB cellsignalfrq 65696c9ef1baf2.65696ca0f11c56.93457589 加水印y77811766No ratings yet
- Cyclic GMP Synthesis, Metabolism, and FunctionDocument353 pagesCyclic GMP Synthesis, Metabolism, and FunctionpawnammalNo ratings yet
- Homo ToxicologyDocument279 pagesHomo ToxicologyRafael MontavaNo ratings yet
- AI Pharma 2 (Edited MCQS)Document11 pagesAI Pharma 2 (Edited MCQS)raneemalnaamnehNo ratings yet
- Biomolecules and Cells NotesDocument81 pagesBiomolecules and Cells NotesLily RobinsonNo ratings yet
- Biochem II Mid Sem 2 MCQs Answered SamiDocument339 pagesBiochem II Mid Sem 2 MCQs Answered Samicarolinegatwiri08No ratings yet
- BIOC1020 Course Document - Outline - SSCDocument4 pagesBIOC1020 Course Document - Outline - SSCbrownjudeaaNo ratings yet
- PharmaDocument147 pagesPharmaDrChauhanNo ratings yet
- Second Messenger SystemsDocument4 pagesSecond Messenger SystemsNatalia Evtikhova EvtikhovNo ratings yet
- Cell SignalingDocument43 pagesCell SignalingRIFAM zNo ratings yet
- Preliminary Term Examination Cytogenetics 001Document16 pagesPreliminary Term Examination Cytogenetics 001MICHAEL JOHN AGUILARNo ratings yet
- Receptor Lecture 1Document24 pagesReceptor Lecture 1Bachelor Gaming CommunityNo ratings yet
- 02 Hormones PDFDocument8 pages02 Hormones PDFKaram SaadNo ratings yet
- BIOM1050: Receptors, Neurons, Synapses & Nervous Systems Cell Communication & Receptor FamiliesDocument12 pagesBIOM1050: Receptors, Neurons, Synapses & Nervous Systems Cell Communication & Receptor FamiliesmichelleNo ratings yet
- Komunikasi Sel PDFDocument64 pagesKomunikasi Sel PDFMuhammad RizaldiNo ratings yet
- Cell SignalingDocument87 pagesCell SignalingZaw Linn PhyoNo ratings yet
- Cell SignalingDocument38 pagesCell SignalingTukai KulkarniNo ratings yet
- Modeling Cellular Signaling Systems: An Abstraction-Refinement ApproachDocument5 pagesModeling Cellular Signaling Systems: An Abstraction-Refinement ApproachdhermitNo ratings yet