Download as pdf or txt
You might also like
- Reassortant Strains of Infectious Bursal Disease VirusDocument24 pagesReassortant Strains of Infectious Bursal Disease VirusJovanaNo ratings yet
- Journal Pre-Proof: Antiviral ResearchDocument44 pagesJournal Pre-Proof: Antiviral ResearchMohammed Shuaib AhmedNo ratings yet
- Perspective: Sars-Cov-2 Vaccines: Status ReportDocument7 pagesPerspective: Sars-Cov-2 Vaccines: Status ReportDeborah BasaNo ratings yet
- Whole Genome Sequencing of Porcine Reproductive and Respiratory Syndrome Virus 2 (PRRSV) From Field Clinical Samples Improves The Genomic Surveillance of The VirusDocument36 pagesWhole Genome Sequencing of Porcine Reproductive and Respiratory Syndrome Virus 2 (PRRSV) From Field Clinical Samples Improves The Genomic Surveillance of The VirusJephte LaputNo ratings yet
- Full Genome EvolutionaryDocument4 pagesFull Genome Evolutionarydoctor gardiNo ratings yet
- Immunity Volume 52 Issue 4 2020 (Doi 10.1016 - J.immuni.2020.03.007) Amanat, Fatima Krammer, Florian - SARS-CoV-2 Vaccines - Status ReportDocument7 pagesImmunity Volume 52 Issue 4 2020 (Doi 10.1016 - J.immuni.2020.03.007) Amanat, Fatima Krammer, Florian - SARS-CoV-2 Vaccines - Status ReportShreyasri SainNo ratings yet
- Preprint Not Peer Reviewed: Neospora Caninum Infection in Sheep: A Systematic Review andDocument55 pagesPreprint Not Peer Reviewed: Neospora Caninum Infection in Sheep: A Systematic Review andVinícius De Paula CastroNo ratings yet
- ch13Document52 pagesch13Renata CezarNo ratings yet
- 378 FullDocument10 pages378 FullHedi SusantoNo ratings yet
- 10 1016@j Vetmic 2019 108522Document7 pages10 1016@j Vetmic 2019 108522Nicolas Carrillo GodoyNo ratings yet
- Porcine ParvovirusDocument14 pagesPorcine ParvovirusRaquel Elena Martínez LópezNo ratings yet
- A Non-Haemadsorbing Live-Attenuated Virus Vaccine Candidate Protects Pigs Against Contemporary Pandemic Genotype Ii African Swine Fever VirusDocument29 pagesA Non-Haemadsorbing Live-Attenuated Virus Vaccine Candidate Protects Pigs Against Contemporary Pandemic Genotype Ii African Swine Fever VirusJovanaNo ratings yet
- Full-Genome Evolutionary Analysis of The Novel Corona Virus (2019-nCoV)Document5 pagesFull-Genome Evolutionary Analysis of The Novel Corona Virus (2019-nCoV)sivebi7088No ratings yet
- Biopolym - Cell 2018 34 5 400 enDocument9 pagesBiopolym - Cell 2018 34 5 400 enАнна ШаповаловаNo ratings yet
- Viruses 14Document13 pagesViruses 14IdoNo ratings yet
- Journal of Virology-2010-Graham-3134.fullDocument13 pagesJournal of Virology-2010-Graham-3134.fullJohn WilliamsonNo ratings yet
- 20220308483506v1full 230222 222641Document37 pages20220308483506v1full 230222 222641Made BagiNo ratings yet
- Insights Into Cross-Species Evolution of Novel Human Coronavirus 2019-Ncov and Defining Immune Determinants For Vaccine DevelopmentDocument17 pagesInsights Into Cross-Species Evolution of Novel Human Coronavirus 2019-Ncov and Defining Immune Determinants For Vaccine Developmentkak.isNo ratings yet
- Highly Pathogenic AvianDocument4 pagesHighly Pathogenic Avianلؤي فارسNo ratings yet
- Cryptococcosis: Eileen K. Maziarz,, John R. PerfectDocument28 pagesCryptococcosis: Eileen K. Maziarz,, John R. PerfectMayra Alejandra Prada SerranoNo ratings yet
- 1 AmaanDocument16 pages1 Amaanbrainhub50No ratings yet
- Investigacion Acerca Del MoquilloDocument8 pagesInvestigacion Acerca Del MoquilloAlexandra GuerreroNo ratings yet
- Identification of 2019-Ncov Related Coronaviruses in Malayan Pangolins in Southern ChinaDocument22 pagesIdentification of 2019-Ncov Related Coronaviruses in Malayan Pangolins in Southern ChinaismailNo ratings yet
- Reviewer DairyDocument43 pagesReviewer DairyLandcel Clarence ArcedoNo ratings yet
- AvianPneumovirus APV PDFDocument7 pagesAvianPneumovirus APV PDFRayza Lubis0% (1)
- The Establishment of Reference Sequence For Sars Cov 2 and Variation AnalysisDocument8 pagesThe Establishment of Reference Sequence For Sars Cov 2 and Variation AnalysisYulaimy BatistaNo ratings yet
- Diversidad Genética y Evolución de SARS-CoV-2Document3 pagesDiversidad Genética y Evolución de SARS-CoV-2Jonatan LZNo ratings yet
- Factores Asociados PRRVSDocument11 pagesFactores Asociados PRRVSJuan Camilo Gomez GarciaNo ratings yet
- 2623 FullDocument2 pages2623 FullNefri ArshintaNo ratings yet
- 1 s2.0 S1567134814003645 MainDocument9 pages1 s2.0 S1567134814003645 Maintemiska amatmuNo ratings yet
- Pathogens 09 00829 v2Document19 pagesPathogens 09 00829 v2jhammyilNo ratings yet
- 670109v2 FullDocument23 pages670109v2 Fullbotond77No ratings yet
- 2019-Novel Goose Parvovirus in Domestic Linwu Sheldrakes With Short Beak and Dwarfism Syndrome, ChinaDocument6 pages2019-Novel Goose Parvovirus in Domestic Linwu Sheldrakes With Short Beak and Dwarfism Syndrome, ChinaTrung Kiên LêNo ratings yet
- Sanchez Seco y Cols 2005Document9 pagesSanchez Seco y Cols 2005roger duranNo ratings yet
- Fvets 09 1080397Document11 pagesFvets 09 1080397botond77No ratings yet
- NIH Public Access: Author ManuscriptDocument24 pagesNIH Public Access: Author ManuscriptVipul BhriguNo ratings yet
- A Novel Live DNA Tagging System For African Swine Fever VirusDocument12 pagesA Novel Live DNA Tagging System For African Swine Fever VirusJovanaNo ratings yet
- UntitledDocument26 pagesUntitledLaura SanchezNo ratings yet
- Development of A Duplex Real-Time RT-PCR Assay For The Detection and Identification of Two Subgroups of Human Metapneumovirus in A Single TubeDocument7 pagesDevelopment of A Duplex Real-Time RT-PCR Assay For The Detection and Identification of Two Subgroups of Human Metapneumovirus in A Single Tubejan yerenaNo ratings yet
- MainDocument37 pagesMainCarla CANo ratings yet
- SARS-CoV-2 and Covid19 Lecture 3 - 2Document5 pagesSARS-CoV-2 and Covid19 Lecture 3 - 2Aishwarya SinghNo ratings yet
- Influenza Virus H5N1 EssayDocument19 pagesInfluenza Virus H5N1 EssayrbfaliNo ratings yet
- Genome Wide Analysis of Sars Cov 2 Virus Strains Circulating Worldwide Implicates HeterogeneityDocument9 pagesGenome Wide Analysis of Sars Cov 2 Virus Strains Circulating Worldwide Implicates HeterogeneityazddazdazdazdNo ratings yet
- Silva Severino Etal IAM 2020 COVID-19Document4 pagesSilva Severino Etal IAM 2020 COVID-19raraelNo ratings yet
- Epidemiology of African Swine Fever VirusDocument7 pagesEpidemiology of African Swine Fever Viruslusitania IkengNo ratings yet
- Avian Influenza Viruses in Mammals: L.A. Reperant, G.F. Rimmelzwaan & T. KuikenDocument24 pagesAvian Influenza Viruses in Mammals: L.A. Reperant, G.F. Rimmelzwaan & T. KuikenSergio OblivionNo ratings yet
- 2022, Plant, HyperVirulence, PsV5, Puccinia, Agrobacterium-Mediated Transformation AND Plasmid Transformation by PEG - FusariumDocument14 pages2022, Plant, HyperVirulence, PsV5, Puccinia, Agrobacterium-Mediated Transformation AND Plasmid Transformation by PEG - FusariumGorae DollNo ratings yet
- 705 2016 Article 2871Document16 pages705 2016 Article 2871Jeann LealNo ratings yet
- 6arenaviruses Zoonotic Jay 2005Document12 pages6arenaviruses Zoonotic Jay 2005Aaron SolatorioNo ratings yet
- Infectious Hypodermal and Haematopoietic Necrosis: 1. ScopeDocument18 pagesInfectious Hypodermal and Haematopoietic Necrosis: 1. ScopeandiniNo ratings yet
- Affiliations:: Zlshi@wh - Iov.cnDocument18 pagesAffiliations:: Zlshi@wh - Iov.cnMuraleetharan_BNo ratings yet
- Nucleotide Sequence and of Aleutian Mink Disease Parvovirus (ADV) : Sequence Comparisons Nonpathogenic and Pathogenic Strain of ADVDocument13 pagesNucleotide Sequence and of Aleutian Mink Disease Parvovirus (ADV) : Sequence Comparisons Nonpathogenic and Pathogenic Strain of ADVhappycold00No ratings yet
- What Makes Cryptococcus Neoformans A Pathogen?: Emerging Infectious Diseases March 1998Document14 pagesWhat Makes Cryptococcus Neoformans A Pathogen?: Emerging Infectious Diseases March 1998Dwika AudiyanandaNo ratings yet
- Chikungunya ReviewDocument23 pagesChikungunya ReviewLina RojasNo ratings yet
- 8 ResearchProjectProposalDocument11 pages8 ResearchProjectProposalderrickkashimba240No ratings yet
- Syndrome Du Nez BlancDocument17 pagesSyndrome Du Nez BlancCyrielle ZNo ratings yet
- Adaptation of HIV-1 To Its Human HostDocument8 pagesAdaptation of HIV-1 To Its Human HostImperialAquillaNo ratings yet
- 2020 03 30 015008v1 Full PDFDocument25 pages2020 03 30 015008v1 Full PDFHarto EstoyNo ratings yet
- Middle East Respiratory Syndrome Coronavirus (Mers-Cov) : Announcement of The Coronavirus Study GroupDocument3 pagesMiddle East Respiratory Syndrome Coronavirus (Mers-Cov) : Announcement of The Coronavirus Study GroupBireswar BhattacharyaNo ratings yet
- Green Washing - The Darker Shade of GreenDocument8 pagesGreen Washing - The Darker Shade of GreenJovanaNo ratings yet
- Are Consumers Being GreenwashedDocument1 pageAre Consumers Being GreenwashedJovanaNo ratings yet
- The Great Green Washing MachineDocument31 pagesThe Great Green Washing MachineJovanaNo ratings yet
- PWA CrosswordDocument3 pagesPWA CrosswordJovanaNo ratings yet
- IPSOS Mobile - First - FinalDocument9 pagesIPSOS Mobile - First - FinalJovanaNo ratings yet
- Dr. Shelly Iskandar, SpAkp, SPKJ, M.Si, PHD Webinar PDSKJI Yogya-PfizerDocument55 pagesDr. Shelly Iskandar, SpAkp, SPKJ, M.Si, PHD Webinar PDSKJI Yogya-PfizerKorn IbraNo ratings yet
- Management of ADHD in Children and Young PeopleDocument52 pagesManagement of ADHD in Children and Young PeopleAna Santos0% (1)
- Kertas Soalan Percubaan SPM Bahasa Inggeris Kertas 2 Set A: Sulit 1119/2Document17 pagesKertas Soalan Percubaan SPM Bahasa Inggeris Kertas 2 Set A: Sulit 1119/2Izmeer AshrayNo ratings yet
- Livro - Bariatric Surgery Latest Advances and ProspectsDocument466 pagesLivro - Bariatric Surgery Latest Advances and Prospectsbetozam3No ratings yet
- Cdem 19december WebDocument32 pagesCdem 19december WebRicardo Jonathan Ayala GarciaNo ratings yet
- Intermittent Fasting - Surprising Update - Harvard Health Blog - Harvard Health Publishing1Document14 pagesIntermittent Fasting - Surprising Update - Harvard Health Blog - Harvard Health Publishing1Piter Kiiro100% (1)
- Antenatal Assessment of Fetal Well Being: DR - Abdulkarim Ahmed MohamudDocument46 pagesAntenatal Assessment of Fetal Well Being: DR - Abdulkarim Ahmed MohamudPeterNo ratings yet
- Review of The TheoryDocument8 pagesReview of The TheoryPriskaCliquersNo ratings yet
- (Blue Books of Practical Neurology 27) W. Ian McDonald and John H. Noseworthy (Eds.) - Multiple Sclerosis 2-Academic Press, Elsevier (2003)Document377 pages(Blue Books of Practical Neurology 27) W. Ian McDonald and John H. Noseworthy (Eds.) - Multiple Sclerosis 2-Academic Press, Elsevier (2003)Pedro BarrosNo ratings yet
- Improper Waste Disposal Effects of Publi PDFDocument2 pagesImproper Waste Disposal Effects of Publi PDFlakshayNo ratings yet
- 3 Day Whole Body Football Strength Workout - Muscle & StrengthDocument11 pages3 Day Whole Body Football Strength Workout - Muscle & StrengthTom HochhalterNo ratings yet
- Geo Exocad Library Selection - 2c0c3Document4 pagesGeo Exocad Library Selection - 2c0c3xigon65878No ratings yet
- Cannondale 2012 IQ200 Cycling Computer FINALDocument4 pagesCannondale 2012 IQ200 Cycling Computer FINALAdrian Popescu0% (1)
- Speech-Language Pathologist Interventions For Communication in Moderate-Severe Dementia - A Systematic ReviewDocument17 pagesSpeech-Language Pathologist Interventions For Communication in Moderate-Severe Dementia - A Systematic ReviewCarol TibaduizaNo ratings yet
- Self - EsteemDocument22 pagesSelf - EsteemNirmal RajNo ratings yet
- Yoga and PainDocument29 pagesYoga and PainGanesh Babu100% (1)
- Collocations IELTSDocument4 pagesCollocations IELTSinfinityieltsjobNo ratings yet
- MCN NCPDocument4 pagesMCN NCPPEARL CHRISTINE CUDALNo ratings yet
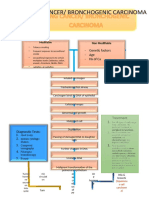
- Lung Cancer PathophysiologyDocument7 pagesLung Cancer PathophysiologyKristian Karl Bautista Kiw-isNo ratings yet
- FM8-35 Transportation of The Sick and Wounded 1970Document211 pagesFM8-35 Transportation of The Sick and Wounded 1970dieudecafeNo ratings yet
- Program DesignDocument19 pagesProgram DesignAmogh PanditNo ratings yet
- ĐỀ 26.MH2022.HSDocument7 pagesĐỀ 26.MH2022.HSThao Thanh PhamNo ratings yet
- Hubungan Antara Pola Makan Dan Status Gizi Dengan Tingkat Kebugaran Atlet DayungDocument7 pagesHubungan Antara Pola Makan Dan Status Gizi Dengan Tingkat Kebugaran Atlet Dayunggaming indoNo ratings yet
- 05 20 Jane LewisDocument34 pages05 20 Jane LewisPrince G.No ratings yet
- Vge ProfileDocument13 pagesVge ProfilexsupermanNo ratings yet
- Ielts ReadingDocument10 pagesIelts Readingruby Javaid21No ratings yet
- Capstone ProjectDocument61 pagesCapstone ProjectShweta GangurdeNo ratings yet
- American College of Physicians Ethics Manual: Annals of Internal MedicineDocument32 pagesAmerican College of Physicians Ethics Manual: Annals of Internal MedicineRitika AgarwalNo ratings yet
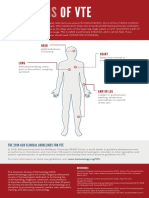
- Signs of VTE Infographic PDFDocument1 pageSigns of VTE Infographic PDFJulia Martha LinardiNo ratings yet
- Economy by Studyshed Jdp5eqDocument44 pagesEconomy by Studyshed Jdp5eqVijaya Prabhu R100% (1)