Download as pdf or txt
You might also like
- Common Diseases of Companion Animals, 4th EditionDocument609 pagesCommon Diseases of Companion Animals, 4th EditionEnzo VicencioNo ratings yet
- First Aid Assess-Answer SheetDocument9 pagesFirst Aid Assess-Answer Sheettsrattan8116No ratings yet
- Procedure Antenatal Assessment - 1Document6 pagesProcedure Antenatal Assessment - 1Kinjal Vasava92% (12)
- Smart Exercise - Burning Fat, Getting Fit - 1994Document157 pagesSmart Exercise - Burning Fat, Getting Fit - 1994Anonymous cCq53ry9100% (3)
- NCP FORM For TetralogyDocument3 pagesNCP FORM For TetralogyGraceMelendres100% (3)
- Articulo 1 PDFDocument8 pagesArticulo 1 PDFJhuliana CabreraNo ratings yet
- Cardiac Electrophysiol Clin 2021 13 4 711-20Document10 pagesCardiac Electrophysiol Clin 2021 13 4 711-20Biblioteca Centro Médico De Mar del PlataNo ratings yet
- Junctional Ectopic Tachycardia in Infants and Children: Ranjit I. Kylat MD - Ricardo A. Samson MDDocument8 pagesJunctional Ectopic Tachycardia in Infants and Children: Ranjit I. Kylat MD - Ricardo A. Samson MDCatherine MorrisNo ratings yet
- AVMinthedifferantialdiagnosisof PalatalswellingDocument5 pagesAVMinthedifferantialdiagnosisof PalatalswellingWafya RamschieNo ratings yet
- Managing Systemic Venous Occlusions in Children: Reviewarticle Open AccessDocument11 pagesManaging Systemic Venous Occlusions in Children: Reviewarticle Open AccessakshayajainaNo ratings yet
- Case Reports Intravascular Ultrasound Identification of Intraluminal Embolic Plaque Material During Carotid Angioplasty With StentingDocument5 pagesCase Reports Intravascular Ultrasound Identification of Intraluminal Embolic Plaque Material During Carotid Angioplasty With StentingRicardo Novoa AlvarezNo ratings yet
- WJR 5 143Document13 pagesWJR 5 143andre halimNo ratings yet
- The Cerebral Arterio-Venous Malformations Radiological PortrayalDocument4 pagesThe Cerebral Arterio-Venous Malformations Radiological PortrayalBRNSS Publication Hub InfoNo ratings yet
- Coronary Sinus Diverticulum As A Cause of Resistant Posteroseptal AP AblationDocument4 pagesCoronary Sinus Diverticulum As A Cause of Resistant Posteroseptal AP AblationFaisal QadirNo ratings yet
- Cureus 0010 00000002642 PDFDocument6 pagesCureus 0010 00000002642 PDFMuhammad Nur Ardhi LahabuNo ratings yet
- Seminar: Daniel J Penny, G Wesley Vick IIIDocument10 pagesSeminar: Daniel J Penny, G Wesley Vick IIIMuhammadAldoGiansyahNo ratings yet
- Recognizing Life-Threatening Causes of Syncope 2013Document16 pagesRecognizing Life-Threatening Causes of Syncope 2013Franco Lozano RamírezNo ratings yet
- Aas AhaDocument12 pagesAas AhaArick Frendi AndriyanNo ratings yet
- February 2024 Stroke HighlightsDocument1 pageFebruary 2024 Stroke HighlightsAbdul AzeezNo ratings yet
- Sites of Successful Ventricular Fibrillation Ablation in Bileaflet Mitral Valve Prolapse SyndromeDocument11 pagesSites of Successful Ventricular Fibrillation Ablation in Bileaflet Mitral Valve Prolapse SyndromeDiego Xavier Chango AzanzaNo ratings yet
- International Consensus Statement On Nomenclature and Classification of The Congenital BAV and Its Aortopathy - 2021Document17 pagesInternational Consensus Statement On Nomenclature and Classification of The Congenital BAV and Its Aortopathy - 2021reymistery10No ratings yet
- Ooa90020 543 547Document5 pagesOoa90020 543 547Sugi ArthaNo ratings yet
- IMP Shock Etiol 5y Outcome VA-ECMO Edit 2023Document3 pagesIMP Shock Etiol 5y Outcome VA-ECMO Edit 2023vladbvs16No ratings yet
- Left Atrial Appendage Occlusion 1Document37 pagesLeft Atrial Appendage Occlusion 1Antonia AdinaNo ratings yet
- Pacing Clinical Electrophis - 2022 - MartiniDocument10 pagesPacing Clinical Electrophis - 2022 - MartiniRobertoNo ratings yet
- Compression Fractures Detection On CTDocument8 pagesCompression Fractures Detection On CTAde Cahyo IslamiNo ratings yet
- Compression Fractures Detection On CTDocument8 pagesCompression Fractures Detection On CTAde Cahyo IslamiNo ratings yet
- Cuoco 2017Document8 pagesCuoco 2017vermashrey2122No ratings yet
- NIH Public Access: Mechanisms of DefibrillationDocument30 pagesNIH Public Access: Mechanisms of DefibrillationIrfan AchmadNo ratings yet
- Incidence of Extrinsic Compression of The Internal Jugular Vein in Unselected Patients Undergoing CT AngiographyDocument4 pagesIncidence of Extrinsic Compression of The Internal Jugular Vein in Unselected Patients Undergoing CT AngiographySuraj DubeyNo ratings yet
- Q Wave in MI PDFDocument9 pagesQ Wave in MI PDFBasant GyawaliNo ratings yet
- Aneurysmal Bone Cyst: Review ArticleDocument9 pagesAneurysmal Bone Cyst: Review ArticleJeyachandran MariappanNo ratings yet
- Intracranial Dural Arteriovenous Fistulae: Presentation and Natural History ClassificationDocument3 pagesIntracranial Dural Arteriovenous Fistulae: Presentation and Natural History ClassificationFinger JempolNo ratings yet
- Supratentorial Arteriovenous MalformationsDocument4 pagesSupratentorial Arteriovenous MalformationsMorteza Mazloom Farsi BafNo ratings yet
- ++veterinary Surgery - 2023 - Banks - Influence of Extreme Brachycephalic Conformation On Perioperative ComplicationsDocument13 pages++veterinary Surgery - 2023 - Banks - Influence of Extreme Brachycephalic Conformation On Perioperative ComplicationsŞahin ŞahinNo ratings yet
- (10920684 - Neurosurgical Focus) Arteriovenous Malformation Presenting With Epilepsy - A Multimodal Approach To Diagnosis and TreatmentDocument7 pages(10920684 - Neurosurgical Focus) Arteriovenous Malformation Presenting With Epilepsy - A Multimodal Approach To Diagnosis and TreatmentNurul FajriNo ratings yet
- Arritmia SVDocument17 pagesArritmia SVCésar Lobo TarangoNo ratings yet
- Corpus Callosotomy With Disconnection SyndromeDocument8 pagesCorpus Callosotomy With Disconnection SyndromeRavi ChandraNo ratings yet
- Wolff-Parkinson-White Syndrome: Anatomy, Epidemiology, Clinical Manifestations, and Diagnosis - UpToDateDocument46 pagesWolff-Parkinson-White Syndrome: Anatomy, Epidemiology, Clinical Manifestations, and Diagnosis - UpToDateKarla ChongNo ratings yet
- Livingston 2017Document7 pagesLivingston 2017Hernan Dario Florez RojasNo ratings yet
- Nuclear Cardiology: Role in The World of Multimodality Cardiac ImagingDocument5 pagesNuclear Cardiology: Role in The World of Multimodality Cardiac ImagingElena FlorentinaNo ratings yet
- Neuroimaging of Spinal Canal StenosisDocument17 pagesNeuroimaging of Spinal Canal Stenosisrulitoss_41739No ratings yet
- Sonopathology 1Document9 pagesSonopathology 1Ivette M. Carroll D.No ratings yet
- Maus 2012Document29 pagesMaus 2012211097salNo ratings yet
- Arterialinjuryinthe Upperextremity: Evaluation, Strategies, and Anticoagulation ManagementDocument11 pagesArterialinjuryinthe Upperextremity: Evaluation, Strategies, and Anticoagulation Managementcirugia plastica uisNo ratings yet
- Discussion of DD of BTLDocument8 pagesDiscussion of DD of BTLshofidhiaaaNo ratings yet
- Wolff-Parkinson-White Syndrome Lessons Learnt and Lessons RemainingDocument6 pagesWolff-Parkinson-White Syndrome Lessons Learnt and Lessons RemainingKarla ChongNo ratings yet
- 01 Cir 53 1 80 PDFDocument6 pages01 Cir 53 1 80 PDFsheilaNo ratings yet
- Arterial Clamping Leads To Stenosis at Clamp Sites After Femoropopliteal Bypass SurgeryDocument9 pagesArterial Clamping Leads To Stenosis at Clamp Sites After Femoropopliteal Bypass SurgeryAyhan ComertNo ratings yet
- Orbital AVMDocument7 pagesOrbital AVMDedeh KurniasihNo ratings yet
- Cystic Lesions of The Brain A Classification Based On PathogenesisDocument16 pagesCystic Lesions of The Brain A Classification Based On Pathogenesisveerraju tvNo ratings yet
- Wong 2020Document5 pagesWong 2020samNo ratings yet
- Acute and Episodic Vestibular Syndromes Caused by Ischemic Stroke: Predilection Sites and Risk FactorsDocument12 pagesAcute and Episodic Vestibular Syndromes Caused by Ischemic Stroke: Predilection Sites and Risk FactorsKarl Martin PinedaNo ratings yet
- Brachial Plexus Birth Palsy: An Overview of Early Treatment ConsiderationsDocument7 pagesBrachial Plexus Birth Palsy: An Overview of Early Treatment Considerationsphannghia.ydsNo ratings yet
- Anorectal Malformation: The Etiological FactorsDocument10 pagesAnorectal Malformation: The Etiological FactorsHasri Rina WalastriNo ratings yet
- Brain Arteriovenous MalformationsDocument20 pagesBrain Arteriovenous MalformationsTony NgNo ratings yet
- Characterization of Congenital Anomalies in Individuals With Choanal AtresiaDocument5 pagesCharacterization of Congenital Anomalies in Individuals With Choanal AtresiaAndi SanapatiNo ratings yet
- TV Dreapta Vs StangaDocument15 pagesTV Dreapta Vs StangaStefana MoisaNo ratings yet
- The Features of ECG and Echocg Functional Parameters in Children With Congenital Heart Disease in The Postoperative PeriodDocument5 pagesThe Features of ECG and Echocg Functional Parameters in Children With Congenital Heart Disease in The Postoperative PeriodCentral Asian StudiesNo ratings yet
- Highlights October-2023-Stroke-HighlightsDocument1 pageHighlights October-2023-Stroke-HighlightsAbdul AzeezNo ratings yet
- Pathogenesis, Evaluation, and ManagementDocument13 pagesPathogenesis, Evaluation, and Managementshentujin1997No ratings yet
- Advances in Cardiovascular ImagingDocument9 pagesAdvances in Cardiovascular ImagingAlina SarbuNo ratings yet
- Decoding Cardiac Electrophysiology: Understanding the Techniques and Defining the JargonFrom EverandDecoding Cardiac Electrophysiology: Understanding the Techniques and Defining the JargonAfzal SohaibNo ratings yet
- Clinical Handbook of Cardiac ElectrophysiologyFrom EverandClinical Handbook of Cardiac ElectrophysiologyBenedict M. GloverNo ratings yet
- Molecular Basis of Drug Pesticide Action: Neurotox '91 &Document370 pagesMolecular Basis of Drug Pesticide Action: Neurotox '91 &Ricardo QuinteiroNo ratings yet
- Electrocardiography HistoryDocument115 pagesElectrocardiography HistoryRicardo QuinteiroNo ratings yet
- Torbal 2004Document9 pagesTorbal 2004Ricardo QuinteiroNo ratings yet
- Ion Channels Family OverviewDocument3 pagesIon Channels Family OverviewRicardo QuinteiroNo ratings yet
- Brown 2011Document6 pagesBrown 2011Ricardo QuinteiroNo ratings yet
- Electrotonic CurrentDocument5 pagesElectrotonic CurrentRicardo QuinteiroNo ratings yet
- Cardiac Sodium Channelopathies: Ion Channels, Receptors and TransportersDocument15 pagesCardiac Sodium Channelopathies: Ion Channels, Receptors and TransportersRicardo QuinteiroNo ratings yet
- Ion ChannelsDocument3 pagesIon ChannelsRicardo QuinteiroNo ratings yet
- Two Dominant Patterns of Low Anterior Resection Syndrome and Their Effects On Patients' Quality of LifeDocument11 pagesTwo Dominant Patterns of Low Anterior Resection Syndrome and Their Effects On Patients' Quality of LifeRicardo QuinteiroNo ratings yet
- Colorectal Disease - 2021 - Christensen - Management Guidelines For Low Anterior Resection Syndrome The MANUEL ProjectDocument15 pagesColorectal Disease - 2021 - Christensen - Management Guidelines For Low Anterior Resection Syndrome The MANUEL ProjectRicardo QuinteiroNo ratings yet
- Ryanodine Receptor Channelopathies: Invited ReviewDocument14 pagesRyanodine Receptor Channelopathies: Invited ReviewRicardo QuinteiroNo ratings yet
- Sakr 2020Document10 pagesSakr 2020Ricardo QuinteiroNo ratings yet
- K 7 Channelopathies: Ion Channels, Receptors and TransportersDocument12 pagesK 7 Channelopathies: Ion Channels, Receptors and TransportersRicardo QuinteiroNo ratings yet
- Single-Nucleotide Polymorphisms in Vascular Ca - Activated K - Channel Genes and Cardiovascular DiseaseDocument9 pagesSingle-Nucleotide Polymorphisms in Vascular Ca - Activated K - Channel Genes and Cardiovascular DiseaseRicardo QuinteiroNo ratings yet
- HCN-related Channelopathies: Invited ReviewDocument11 pagesHCN-related Channelopathies: Invited ReviewRicardo QuinteiroNo ratings yet
- MODULE 2 (Chapter 2.1)Document12 pagesMODULE 2 (Chapter 2.1)Malapitan, Kiana Blanca A.No ratings yet
- Fuchs S Et Al. Pediatrics. 2016 138Document10 pagesFuchs S Et Al. Pediatrics. 2016 138Esteban RamosNo ratings yet
- MAP of NDPQ QuestionsDocument5 pagesMAP of NDPQ QuestionsHenri IndrawanNo ratings yet
- Heart MurmursDocument18 pagesHeart MurmursRobby Wiranata WijayaNo ratings yet
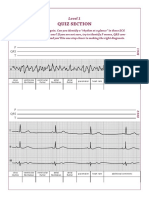
- Quiz Section: Level 1Document4 pagesQuiz Section: Level 1AinunZamiraNo ratings yet
- Euad 043Document29 pagesEuad 043Pedro SeguraNo ratings yet
- Echocardiogram MayoDocument4 pagesEchocardiogram MayoMarta KitaNo ratings yet
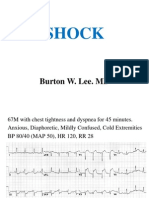
- Cardiogenic ShockDocument31 pagesCardiogenic ShockAbdifatah OomaneNo ratings yet
- Vital Sign PresentationDocument94 pagesVital Sign PresentationlovekovinNo ratings yet
- Stemcell CardiovascularDocument29 pagesStemcell CardiovascularAdi WijayantoNo ratings yet
- Heart Disease in PregnancyDocument21 pagesHeart Disease in PregnancyS VaibhavNo ratings yet
- CONCEPT - QuestionsDocument2 pagesCONCEPT - Questionsgreen_archerNo ratings yet
- Drugs Used in Cardiac Failure (CCF)Document8 pagesDrugs Used in Cardiac Failure (CCF)Faria Islam Juhi100% (1)
- The Six Second ECGDocument6 pagesThe Six Second ECGRaquel GirónNo ratings yet
- Hemodinamic Description of Volleyball Atlet Before and After Competing in Banyumas DistrictDocument11 pagesHemodinamic Description of Volleyball Atlet Before and After Competing in Banyumas DistrictariNo ratings yet
- Cristina Barsolo VS SSSDocument2 pagesCristina Barsolo VS SSSHasmer Maulana AmalNo ratings yet
- Connective Tissue Disorders PhysicalDocument37 pagesConnective Tissue Disorders PhysicalR DNo ratings yet
- What Is Tetralogy of FallotDocument2 pagesWhat Is Tetralogy of FallotJamaica Cruz San Pedro100% (1)
- MyocarditisDocument29 pagesMyocarditishastonniiNo ratings yet
- Soal Reading B.ing 22 Feb (Najhan)Document9 pagesSoal Reading B.ing 22 Feb (Najhan)Green AlonsoNo ratings yet
- Notes of CH 6 Life Process Part-II - Class 10th Science Study RankersDocument7 pagesNotes of CH 6 Life Process Part-II - Class 10th Science Study RankerssamyNo ratings yet
- January 2022 (IAL) QPDocument28 pagesJanuary 2022 (IAL) QPMaria Shabbir PanjiwalaNo ratings yet
- PDA Case StudyDocument5 pagesPDA Case StudyDiana Andrea AyongaoNo ratings yet
- Work Physiology: "Underloading", in Which Human Capacities AreDocument35 pagesWork Physiology: "Underloading", in Which Human Capacities AreLance SantiagoNo ratings yet
- ICD - Implantable Cardioverter DefibrillatorDocument8 pagesICD - Implantable Cardioverter DefibrillatorJon SmithNo ratings yet