Download as pdf or txt
You might also like
- AACR 2022 Proceedings: Part A Online-Only and April 10From EverandAACR 2022 Proceedings: Part A Online-Only and April 10No ratings yet
- Transition Care PlanDocument5 pagesTransition Care PlangyanendraNo ratings yet
- Label KuningDocument2,405 pagesLabel KuningAll Malik Ha100% (5)
- Determination of Anticancer Potential of A Novel Pharmacolo 2022 Saudi PharmDocument10 pagesDetermination of Anticancer Potential of A Novel Pharmacolo 2022 Saudi Pharmcfy28722psNo ratings yet
- A Comprehensive Review On The Indazole Based Derivatives As Targeted Anticancer AgentsDocument26 pagesA Comprehensive Review On The Indazole Based Derivatives As Targeted Anticancer AgentsAlexandra BurlacuNo ratings yet
- 1 s2.0 S1319562X23000542 MainDocument10 pages1 s2.0 S1319562X23000542 Mainmonu kashyapNo ratings yet
- Bladder Cancer Cell Viability Inhibition and Apoptosis I - 2018 - Saudi JournalDocument5 pagesBladder Cancer Cell Viability Inhibition and Apoptosis I - 2018 - Saudi JournalErick Dario P ChNo ratings yet
- Research Gate-Health Benifits of Cor-MDocument7 pagesResearch Gate-Health Benifits of Cor-MHarendra ChunduruNo ratings yet
- Algae A Robust Living Material Against CancerDocument23 pagesAlgae A Robust Living Material Against CancerJuan David Rodriguez HurtadoNo ratings yet
- Synthesis, Anticancer and Antioxidant Properties of New Indole and Pyranoindole DerivativesDocument13 pagesSynthesis, Anticancer and Antioxidant Properties of New Indole and Pyranoindole DerivativesCesar EscareñoNo ratings yet
- NO. 12. Lee CH. 2019-Reactive Oxygen Species Mediate The Chemopreventive Effects of Syringin in Breast CancerDocument12 pagesNO. 12. Lee CH. 2019-Reactive Oxygen Species Mediate The Chemopreventive Effects of Syringin in Breast CancerVincent AriesNo ratings yet
- Breast CancerhggDocument20 pagesBreast CancerhggCatherine RajanNo ratings yet
- 10 1016@j Biopha 2019 109142Document13 pages10 1016@j Biopha 2019 109142CoNo ratings yet
- Nuevo Forma de Investigar El Cancer PancreaticoDocument8 pagesNuevo Forma de Investigar El Cancer PancreaticovillalobosgarzacarlosfranciscoNo ratings yet
- Cordyceps Nakamura2015Document4 pagesCordyceps Nakamura2015Jorge Luis Plasencia CubaNo ratings yet
- Herbal and Synthetic Drug Combinations in Cancer Therapy A ReviewDocument6 pagesHerbal and Synthetic Drug Combinations in Cancer Therapy A ReviewEditor IJTSRDNo ratings yet
- The Anticancer Activity of Bioactive Compounds From Medicinal Plants: An ReviewDocument7 pagesThe Anticancer Activity of Bioactive Compounds From Medicinal Plants: An ReviewVinayNo ratings yet
- 1 s2.0 S187853522100068X MainDocument18 pages1 s2.0 S187853522100068X MainsonNo ratings yet
- Cancers 14 04299Document15 pagesCancers 14 04299Gabu BuitronNo ratings yet
- Sabira Sultana REVIEWapjcpDocument11 pagesSabira Sultana REVIEWapjcpDiana RomeroNo ratings yet
- Alcaloids and CancerDocument14 pagesAlcaloids and CancerMed ZinNo ratings yet
- Cannabis BiomoleculesDocument11 pagesCannabis BiomoleculesWallfayerNo ratings yet
- Grzyb I Rak2Document11 pagesGrzyb I Rak2Matt NeputinNo ratings yet
- Evaluation of Biogenically Synthesized MgO NPsDocument8 pagesEvaluation of Biogenically Synthesized MgO NPstobyNo ratings yet
- Arun Et Al 2017 Anti Colon Cancer Activity of M. KoenigiiDocument12 pagesArun Et Al 2017 Anti Colon Cancer Activity of M. KoenigiiRavi P ShaliwalNo ratings yet
- Turmeric Is Therapeutic in Vivo On Patient-Derived Colorectal Cancer Xenografts: Inhibition of Growth, Metastasis, and Tumor RecurrenceDocument16 pagesTurmeric Is Therapeutic in Vivo On Patient-Derived Colorectal Cancer Xenografts: Inhibition of Growth, Metastasis, and Tumor RecurrenceFilipe GonçalvesNo ratings yet
- Anti-Cancer Potential of Sophoridine and Its Derivatives Recent Progress and Future PerspectivesDocument20 pagesAnti-Cancer Potential of Sophoridine and Its Derivatives Recent Progress and Future PerspectivesHaroon Ur RashidNo ratings yet
- Anticancer Activity of Bee Venom Components Against Breast CancerDocument11 pagesAnticancer Activity of Bee Venom Components Against Breast CancerSergio B.No ratings yet
- Biomaterials: Amit Sharma, Eun-Joong Kim, Hu Shi, Jin Yong Lee, Bong Geun Chung, Jong Seung KimDocument7 pagesBiomaterials: Amit Sharma, Eun-Joong Kim, Hu Shi, Jin Yong Lee, Bong Geun Chung, Jong Seung KimVijay LouNo ratings yet
- Piperine Cancer and SenescenceDocument10 pagesPiperine Cancer and SenescencevinfiftenNo ratings yet
- Venom-Based Peptide Therapy Insights Into Anti-Cancer MechanismDocument23 pagesVenom-Based Peptide Therapy Insights Into Anti-Cancer MechanismMaleny HernándezNo ratings yet
- Wang 2021Document7 pagesWang 2021Portgas D AceNo ratings yet
- Effects of Current Chemotherapy Drugs and Natural Agents in Treating Non-Small Cell Lung CancerDocument5 pagesEffects of Current Chemotherapy Drugs and Natural Agents in Treating Non-Small Cell Lung CancerResearch ParkNo ratings yet
- Jiang, Z., Chen, K., Cheng, L., Yan, B., Qian, W., Cao, J., ... Yang, W. (2017) - Resveratrol y Tratamiento Del Cáncer - Actualizaciones.Document11 pagesJiang, Z., Chen, K., Cheng, L., Yan, B., Qian, W., Cao, J., ... Yang, W. (2017) - Resveratrol y Tratamiento Del Cáncer - Actualizaciones.Luis ReyNo ratings yet
- Vallery A - 23175188a - T3Document4 pagesVallery A - 23175188a - T3niaNo ratings yet
- The Anticancer Properties of Cordycepin and Their Underlying MechanismsDocument11 pagesThe Anticancer Properties of Cordycepin and Their Underlying MechanismsJorge Luis Plasencia CubaNo ratings yet
- Nanoparticles Magnetic CancerDocument8 pagesNanoparticles Magnetic CancerAndres VaronNo ratings yet
- Botany ProjectDocument25 pagesBotany ProjectSunil ShekharNo ratings yet
- Research Article in Vivo Anticancer Activity of Basella Alba Leaf and SeedDocument11 pagesResearch Article in Vivo Anticancer Activity of Basella Alba Leaf and SeedingelitaNo ratings yet
- Referensi Ic50 1Document11 pagesReferensi Ic50 1Detria Rahma GastiNo ratings yet
- Project Breast CancerDocument21 pagesProject Breast CancerDr. Avnish UpadhyayNo ratings yet
- Biology-Driven Cancer Drug Development: Back To The Future: ReviewDocument12 pagesBiology-Driven Cancer Drug Development: Back To The Future: ReviewangelicdispositionNo ratings yet
- 1 Vol. 3 Issue 2 Feb 2016 IJP RE 1810Document12 pages1 Vol. 3 Issue 2 Feb 2016 IJP RE 1810Josimar da RosaNo ratings yet
- 1 s2.0 S0300483X20301670 MainDocument12 pages1 s2.0 S0300483X20301670 MainAhmad SolihinNo ratings yet
- Aspirin and CancerDocument7 pagesAspirin and CancermanoloeNo ratings yet
- Carcinogenesis Failure of Resolution of InflammatiDocument38 pagesCarcinogenesis Failure of Resolution of InflammatiThe Researcher ArgentinaNo ratings yet
- Cordyceps and Medicine Past Present andDocument4 pagesCordyceps and Medicine Past Present andChris GuarascioNo ratings yet
- 1443-Article Text-3899-1-10-20170514Document8 pages1443-Article Text-3899-1-10-20170514Akademisi Bisnis indonesiaNo ratings yet
- Potential of Natural Products in Osteosarcoma TreatmentDocument21 pagesPotential of Natural Products in Osteosarcoma TreatmentOstazNo ratings yet
- Cancer_drug_delivery_in_the_nano_era_An_overview_aDocument15 pagesCancer_drug_delivery_in_the_nano_era_An_overview_aAngelina Diva AnggreiniNo ratings yet
- Iman Et Al Girinimbine As Angiogensis InhibitorDocument12 pagesIman Et Al Girinimbine As Angiogensis InhibitorRavi P ShaliwalNo ratings yet
- Recent Advances in The Development Of.1Document6 pagesRecent Advances in The Development Of.1Prince PatilNo ratings yet
- Biomedicines 11 01506Document25 pagesBiomedicines 11 01506erdogansachirNo ratings yet
- Doxorubicin 2Document10 pagesDoxorubicin 2Afrian RahmandaNo ratings yet
- Saudi Journal of Biological Sciences: Omaima NasirDocument8 pagesSaudi Journal of Biological Sciences: Omaima NasirlanilauNo ratings yet
- Angelica Sinensis Polysaccharide Inhibits Inflammation of Co - 2023 - Arabian JoDocument9 pagesAngelica Sinensis Polysaccharide Inhibits Inflammation of Co - 2023 - Arabian JoLaboratorium BCPekanbaruNo ratings yet
- Ol 18 1 487 PDFDocument12 pagesOl 18 1 487 PDFCasiana GuiNo ratings yet
- Artigo Cisplatina 1Document12 pagesArtigo Cisplatina 1Daiane RochaNo ratings yet
- 5-Azacytidine Prevents Cisplatin Induced Nephrotoxicity and PotentiatesDocument9 pages5-Azacytidine Prevents Cisplatin Induced Nephrotoxicity and PotentiatesTorres Pineda OsvaldoNo ratings yet
- Materials 14 03368Document17 pagesMaterials 14 03368najem AlsalehNo ratings yet
- Plant Derived Extracts CancerDocument38 pagesPlant Derived Extracts CancerAman KaurNo ratings yet
- Regulación de La Resistencia A La Insulina Por Griffola FrondosaDocument13 pagesRegulación de La Resistencia A La Insulina Por Griffola FrondosaPauly Campos OyarzúnNo ratings yet
- Efecto Regulador de Griffola FrondosaDocument11 pagesEfecto Regulador de Griffola FrondosaPauly Campos OyarzúnNo ratings yet
- Hongos Como Fuente de Ácidos GrasosDocument16 pagesHongos Como Fuente de Ácidos GrasosPauly Campos OyarzúnNo ratings yet
- Los Hongos Contienen Genes Asociados A La Biosíntesis de FlavonoidesDocument1 pageLos Hongos Contienen Genes Asociados A La Biosíntesis de FlavonoidesPauly Campos OyarzúnNo ratings yet
- Aplicaciones de Productos de Hongos en Alimentos y Salud HumanaDocument14 pagesAplicaciones de Productos de Hongos en Alimentos y Salud HumanaPauly Campos OyarzúnNo ratings yet
- Hydrophobin Gene Deletion and Environmental Growth Conditions Impact Mechanical Properties of Mycelium by Affecting The Density of The MateriaDocument7 pagesHydrophobin Gene Deletion and Environmental Growth Conditions Impact Mechanical Properties of Mycelium by Affecting The Density of The MateriaPauly Campos OyarzúnNo ratings yet
- Actividad Antioxidante de CoprinusDocument8 pagesActividad Antioxidante de CoprinusPauly Campos OyarzúnNo ratings yet
- Advanced Materials From Fungal Mycelium: Fabrication and Tuning of Physical PropertiesDocument11 pagesAdvanced Materials From Fungal Mycelium: Fabrication and Tuning of Physical PropertiesPauly Campos OyarzúnNo ratings yet
- Effect of Lentinus Edodes Polysaccharide On Oxidative Stress, Immunity Activity and Oral Ulceration of Rats Stimulated by PhenolDocument4 pagesEffect of Lentinus Edodes Polysaccharide On Oxidative Stress, Immunity Activity and Oral Ulceration of Rats Stimulated by PhenolPauly Campos OyarzúnNo ratings yet
- Microbial NanobionicsDocument332 pagesMicrobial NanobionicsPauly Campos OyarzúnNo ratings yet
- Updates Ontropical Mushrooms 2018Document229 pagesUpdates Ontropical Mushrooms 2018Pauly Campos OyarzúnNo ratings yet
- Capability of Selected Mushrooms To Biodegrade PolyethyleneDocument8 pagesCapability of Selected Mushrooms To Biodegrade PolyethylenePauly Campos OyarzúnNo ratings yet
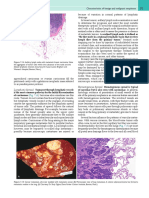
- Histological Classifi Cation of Breast CancerDocument12 pagesHistological Classifi Cation of Breast Cancersilviailie100% (1)
- The New FIGO StagingDocument4 pagesThe New FIGO StagingDr.MohanNo ratings yet
- Medical Diagnosis and Pathophysiology: (Be Sure To List Signs and Symptoms) - Please Indicate Your Source of InformationDocument3 pagesMedical Diagnosis and Pathophysiology: (Be Sure To List Signs and Symptoms) - Please Indicate Your Source of InformationKenneth Smith IINo ratings yet
- Esmo Pocket Guidelines On Breast Cancer 2013 PDFDocument54 pagesEsmo Pocket Guidelines On Breast Cancer 2013 PDFHermawan HmnNo ratings yet
- Neoplasia 2Document57 pagesNeoplasia 2Dyvanal ArifinNo ratings yet
- Test Bank For Maternity and Womens Health Care 11th Edition by LowdermilkDocument11 pagesTest Bank For Maternity and Womens Health Care 11th Edition by Lowdermilkdonnadaviskjqpxitacn100% (32)
- Principles of Surgical OncologyDocument34 pagesPrinciples of Surgical Oncologykaukab azim100% (1)
- Neoplasma CodingDocument59 pagesNeoplasma CodingMuhammad Syamsul ArifinNo ratings yet
- Discrimination of Metastatic From Acute Osteoporotic Compression Spinal Fractures With MRIDocument9 pagesDiscrimination of Metastatic From Acute Osteoporotic Compression Spinal Fractures With MRIChow Tat SingNo ratings yet
- 5.4 Treatment of Early (Stage I and II) Head and Neck Cancer - The Larynx - UpToDateDocument33 pages5.4 Treatment of Early (Stage I and II) Head and Neck Cancer - The Larynx - UpToDateMarco GornattiNo ratings yet
- 2018 - Early Diagnosis and Detection of Breast Cancer - Milosevic2018Document31 pages2018 - Early Diagnosis and Detection of Breast Cancer - Milosevic2018Adriano R. OrtizNo ratings yet
- Cancer PainDocument52 pagesCancer PainIndraYudhi100% (1)
- Brockstein - Head and Neck CancerDocument387 pagesBrockstein - Head and Neck CancerDr. Christian Dimitrov100% (1)
- J Neuro Oncol 2022Document10 pagesJ Neuro Oncol 2022EvelynNo ratings yet
- Oropharingeal Carcinoma Parvathaneni2019Document14 pagesOropharingeal Carcinoma Parvathaneni2019kikicreNo ratings yet
- Prognostic Markers Compared To CD3 TIL in Locally.83Document7 pagesPrognostic Markers Compared To CD3 TIL in Locally.83Yohana Elisabeth GultomNo ratings yet
- Benign and Malignant TumorsDocument22 pagesBenign and Malignant TumorsmoupriyanagNo ratings yet
- Lung Cancer - NCPDocument2 pagesLung Cancer - NCPChen BernardoNo ratings yet
- Factors Associated With Short-Term Mortality After Surgical Oncologic EmergenciesDocument12 pagesFactors Associated With Short-Term Mortality After Surgical Oncologic EmergenciesErinda SafitriNo ratings yet
- Pathology Mci One Liners MCQDocument21 pagesPathology Mci One Liners MCQadiNo ratings yet
- BioSB 2021 Catalog210x280 062821Document398 pagesBioSB 2021 Catalog210x280 062821NANCY VALENCIANo ratings yet
- Hematogenous Spread. Lymphatic Spread.: 275 Characteristics of Benign and Malignant NeoplasmsDocument4 pagesHematogenous Spread. Lymphatic Spread.: 275 Characteristics of Benign and Malignant NeoplasmsGin TokiNo ratings yet
- Reaserch Content On Breast Cancer and Lesson Plan 1 PDFDocument33 pagesReaserch Content On Breast Cancer and Lesson Plan 1 PDFNabanita KunduNo ratings yet
- Pharmacy Practice-I MCQ: A. Nasal CongestionDocument62 pagesPharmacy Practice-I MCQ: A. Nasal CongestionJoyeeta Tahseen KhanNo ratings yet
- 5 4b Childhood Malignancy Part 2 DR Melanie Victoria G DarDocument7 pages5 4b Childhood Malignancy Part 2 DR Melanie Victoria G DarSamatha SamathaNo ratings yet
- Slides LogisticDocument33 pagesSlides LogisticMohmmad RezaNo ratings yet
- Plasma in Cancer TreatmentDocument360 pagesPlasma in Cancer TreatmentLiz SernaNo ratings yet
- Iqvia Institute Global Oncology Trends 2022 ForwebDocument65 pagesIqvia Institute Global Oncology Trends 2022 ForwebYefinadya MfNo ratings yet