Download as pdf or txt
You might also like
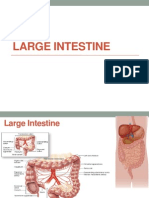
- Large Intestine HistologyDocument24 pagesLarge Intestine HistologyMudassar RoomiNo ratings yet
- Anatomi Genitalia Pria WanitaDocument65 pagesAnatomi Genitalia Pria WanitaTaufik Adi SNo ratings yet
- Gastrointestinal Tract: Intestinal Mucosa, ToxinsDocument9 pagesGastrointestinal Tract: Intestinal Mucosa, ToxinsKhine Kyi ThuNo ratings yet
- The Neonatal Gastrointestinal Tract:: Developmental Anatomy, Physiology, and Clinical ImplicationsDocument9 pagesThe Neonatal Gastrointestinal Tract:: Developmental Anatomy, Physiology, and Clinical ImplicationsegerpratamaNo ratings yet
- Importance of Nutritional Therapy in The Management of Intestinal Diseases: Beyond Energy and Nutrient SupplyDocument12 pagesImportance of Nutritional Therapy in The Management of Intestinal Diseases: Beyond Energy and Nutrient SupplyIsabella María GantivarNo ratings yet
- New Ways of Thinking About (And Teaching About) Intestinal Epithelial FunctionDocument10 pagesNew Ways of Thinking About (And Teaching About) Intestinal Epithelial FunctionSinisa RisticNo ratings yet
- BiomedPap - Bio 201604 0001Document6 pagesBiomedPap - Bio 201604 0001vanebenuzziNo ratings yet
- B1 CellDocument22 pagesB1 CellPierfelice CutrufelliNo ratings yet
- Gut Kidney 1Document10 pagesGut Kidney 1AlfirahmatikaNo ratings yet
- Developmental Microbial Ecology of The Neona 1999 The American Journal of CLDocument11 pagesDevelopmental Microbial Ecology of The Neona 1999 The American Journal of CLJoão PedroNo ratings yet
- Nutrients 12 01185Document18 pagesNutrients 12 01185Zannatun Nahar JhinuNo ratings yet
- Gut Flora in Health and DiseaseDocument8 pagesGut Flora in Health and DiseaseJohnny AtmanNo ratings yet
- Porridge Feeding As Nutritional Management of Typhoid Fever: Take It or Leave It?Document5 pagesPorridge Feeding As Nutritional Management of Typhoid Fever: Take It or Leave It?Nadhira AfifaNo ratings yet
- Pediatrics in Review 1989 DeWitt 6 12Document10 pagesPediatrics in Review 1989 DeWitt 6 12DewinsNo ratings yet
- Impact of Regional Differences Along The Gastrointestinal Tract of HealthyDocument23 pagesImpact of Regional Differences Along The Gastrointestinal Tract of Healthyangiejhoana asprillaNo ratings yet
- Fnut 08 718356Document24 pagesFnut 08 718356Lisa GonçalvesNo ratings yet
- 363 PDF PDFDocument7 pages363 PDF PDFEko Setyo BudiNo ratings yet
- Development of An in Vitro Co-Culture Model To Mimic The Human IntestineDocument13 pagesDevelopment of An in Vitro Co-Culture Model To Mimic The Human IntestineRizki AwaluddinNo ratings yet
- Antenatal CareDocument7 pagesAntenatal CareWegdan AliNo ratings yet
- Diare Non InflamasiDocument4 pagesDiare Non InflamasiSiti MunawwarahNo ratings yet
- Obesity and The Human Microbiome: Ruth E. LeyDocument7 pagesObesity and The Human Microbiome: Ruth E. Leyanka_mihaelaNo ratings yet
- The Mucus and Mucins of The Goblet Cells and EnterocytesDocument23 pagesThe Mucus and Mucins of The Goblet Cells and EnterocytesMacario OtsögaNo ratings yet
- Pediatric Intestinal Failure ReviewDocument12 pagesPediatric Intestinal Failure Reviewvendezu18No ratings yet
- Nutritional Support in Gastrointestinal Diseases: Compromised GutDocument18 pagesNutritional Support in Gastrointestinal Diseases: Compromised GutRoxana CristeaNo ratings yet
- Gi Embryo ComparativeDocument13 pagesGi Embryo ComparativemikeNo ratings yet
- Microbial Activities and Intestinal Homeostasis A Delicate Balance Between Health and DiseaseDocument13 pagesMicrobial Activities and Intestinal Homeostasis A Delicate Balance Between Health and DiseaseitomoralesNo ratings yet
- Saf Frey 2013Document14 pagesSaf Frey 2013Leonardo AzevedoNo ratings yet
- Linking Gut Microbiota and Inflammation To Obesity and Insulin ResistanceDocument11 pagesLinking Gut Microbiota and Inflammation To Obesity and Insulin ResistanceDr LoloNo ratings yet
- CANINE-Pathophysiology and Management of Canine Colonic DiseasesDocument14 pagesCANINE-Pathophysiology and Management of Canine Colonic Diseasestaner_soysuren100% (1)
- Thompson AJ, Et Al.2021Document15 pagesThompson AJ, Et Al.2021Balsam zahharNo ratings yet
- Artis 2008Document10 pagesArtis 2008contramondum1232039No ratings yet
- The Gut-Liver Axis in Liver Disease: Pathophysiological Basis For TherapyDocument20 pagesThe Gut-Liver Axis in Liver Disease: Pathophysiological Basis For TherapyDayane RomanNo ratings yet
- CLN 65p635Document10 pagesCLN 65p635Priady Wira PrasetiaNo ratings yet
- Gut Kidney Cross TalkDocument37 pagesGut Kidney Cross TalkAbdelrahman MokhtarNo ratings yet
- Khoshbin Camilleri 2020 Effects of Dietary Components On Intestinal Permeability in Health and DiseaseDocument20 pagesKhoshbin Camilleri 2020 Effects of Dietary Components On Intestinal Permeability in Health and Diseaserenato cruzNo ratings yet
- Gross Lesions of Alimentary Disease in Adult CattleDocument31 pagesGross Lesions of Alimentary Disease in Adult CattleLukas ZamudioNo ratings yet
- Reviews: You Are What You Eat: Diet, Health and The Gut MicrobiotaDocument22 pagesReviews: You Are What You Eat: Diet, Health and The Gut MicrobiotaHarry YucraNo ratings yet
- Gut-Microbiome-Obesity-And-Metabolic-DysfunctionDocument8 pagesGut-Microbiome-Obesity-And-Metabolic-DysfunctionSandraosoNo ratings yet
- With A Little Help From My Friends-The Role of Microbiota in Thyroid Hormone 2017Document5 pagesWith A Little Help From My Friends-The Role of Microbiota in Thyroid Hormone 2017Aline DantasNo ratings yet
- Gut MicrobiotaDocument17 pagesGut MicrobiotaBarbie mendroliaNo ratings yet
- Laprak Unit Mencit SuzyyDocument13 pagesLaprak Unit Mencit Suzyynanda syafiraNo ratings yet
- Gut Bacteria in Health and DiseaseDocument10 pagesGut Bacteria in Health and DiseasevvvNo ratings yet
- Pnas 201006734Document8 pagesPnas 201006734karimsamihaddad1234No ratings yet
- Cristofolini 2012Document10 pagesCristofolini 2012Ro BellingeriNo ratings yet
- Perubahan Pencernaan Bayi Baru LahirDocument15 pagesPerubahan Pencernaan Bayi Baru LahirrizaNo ratings yet
- GABA ObesityDocument15 pagesGABA ObesityPame RuizNo ratings yet
- Intestinal PermeabilityDocument6 pagesIntestinal PermeabilitysakuraleeshaoranNo ratings yet
- Biology Attachment Yhwh CaresDocument5 pagesBiology Attachment Yhwh CaresanthonybaziraNo ratings yet
- Role of Flavonoids in Intestinal Tight Junction RegulationDocument8 pagesRole of Flavonoids in Intestinal Tight Junction RegulationGina PistolNo ratings yet
- Drug Metabolism in The Malnourished Child: Sarqj MehtaDocument10 pagesDrug Metabolism in The Malnourished Child: Sarqj MehtaIvana SantosoNo ratings yet
- Leptin Signal Transduction Underlies The Differential Metabolic Response of LEW and WKY Rats To Cafeteria DietDocument10 pagesLeptin Signal Transduction Underlies The Differential Metabolic Response of LEW and WKY Rats To Cafeteria Dietjlee_296737No ratings yet
- Jurnal PediatriDocument16 pagesJurnal PediatriRahmat Sayyid ZharfanNo ratings yet
- Complex Interactions of Circadian Rhythms, EatingDocument10 pagesComplex Interactions of Circadian Rhythms, EatingroxxyiNo ratings yet
- Infectious Diarrhea: Cellular and Molecular MechanismsDocument18 pagesInfectious Diarrhea: Cellular and Molecular Mechanismsmedicalfuture2uNo ratings yet
- Role of Barrier Integrity and Dysfunctions in Maintaining The Healthy Gut and Their Health OutcomesDocument15 pagesRole of Barrier Integrity and Dysfunctions in Maintaining The Healthy Gut and Their Health OutcomesGina PistolNo ratings yet
- Gastrointestinal TractDocument3 pagesGastrointestinal TractElgustaNo ratings yet
- Beasleyetal 2015Document13 pagesBeasleyetal 2015Shamsul ArefinNo ratings yet
- Zonulin, A Regulator of Epithelial and Endothelial Barrier FunctionDocument19 pagesZonulin, A Regulator of Epithelial and Endothelial Barrier Functionanna.andreouu321No ratings yet
- Oral Versus Gastrointestinal Mucosal Immune Niches in Homeostasis and AllostasisDocument21 pagesOral Versus Gastrointestinal Mucosal Immune Niches in Homeostasis and Allostasissandip ghoseNo ratings yet
- Nutrients 14 01578Document10 pagesNutrients 14 01578ANDREEA-CORNELIA NECULCEANo ratings yet
- The Flora of The Major Body Systems: The Oral Cavity and The Gastrointestinal TractDocument2 pagesThe Flora of The Major Body Systems: The Oral Cavity and The Gastrointestinal TractAiza Mae Clacio LimNo ratings yet
- Mucosal Membrane Health: The Key to Preventing Inflammatory Conditions, Infections, Toxicity and DegenerationFrom EverandMucosal Membrane Health: The Key to Preventing Inflammatory Conditions, Infections, Toxicity and DegenerationNo ratings yet
- Effects of Long Term Intake of Retinol On Selec 1999 The American Journal ofDocument7 pagesEffects of Long Term Intake of Retinol On Selec 1999 The American Journal ofJoão PedroNo ratings yet
- Vitamin D Supplementation 25 Hydroxyvitamin 1999 The American Journal of CDocument15 pagesVitamin D Supplementation 25 Hydroxyvitamin 1999 The American Journal of CJoão PedroNo ratings yet
- Erratum - 1999 - The American Journal of Clinical NutritionDocument2 pagesErratum - 1999 - The American Journal of Clinical NutritionJoão PedroNo ratings yet
- Energy Density But Not Fat Content of Foods Affe - 1999 - The American Journal oDocument9 pagesEnergy Density But Not Fat Content of Foods Affe - 1999 - The American Journal oJoão PedroNo ratings yet
- Probiotics Prebiotics and Synbiotics Approache 1999 The American JournalDocument6 pagesProbiotics Prebiotics and Synbiotics Approache 1999 The American JournalJoão PedroNo ratings yet
- Lipoprotein Protocols Edited by Jose M Ordovas 199 - 1999 - The American JournDocument2 pagesLipoprotein Protocols Edited by Jose M Ordovas 199 - 1999 - The American JournJoão PedroNo ratings yet
- Whole Body Protein Turnover and Resting Energy e - 1999 - The American Journal oDocument6 pagesWhole Body Protein Turnover and Resting Energy e - 1999 - The American Journal oJoão PedroNo ratings yet
- Energy Expenditure and Substrate Utilization in A - 1999 - The American JournalDocument7 pagesEnergy Expenditure and Substrate Utilization in A - 1999 - The American JournalJoão PedroNo ratings yet
- Lessons For Nutritional Science From - 1999 - The American Journal of ClinicalDocument2 pagesLessons For Nutritional Science From - 1999 - The American Journal of ClinicalJoão PedroNo ratings yet
- Dietary Intake of - Linolenic Acid and Risk of F - 1999 - The American Journal oDocument8 pagesDietary Intake of - Linolenic Acid and Risk of F - 1999 - The American Journal oJoão PedroNo ratings yet
- Prof - Abdul Jabbar N. Al-ShammariDocument51 pagesProf - Abdul Jabbar N. Al-Shammariمختبرات ابوسارةNo ratings yet
- UNIT 1 (Anatomy & Physiology)Document11 pagesUNIT 1 (Anatomy & Physiology)Workinesh Kaynabo KambaloNo ratings yet
- Daftar Pustaka Paper Oftalmia Neonatorum (Sudah Digabung) PDFDocument153 pagesDaftar Pustaka Paper Oftalmia Neonatorum (Sudah Digabung) PDFDewi Sartika100% (1)
- Class 9 MT Science Notes TissueDocument12 pagesClass 9 MT Science Notes TissueKhushi Agrawal100% (1)
- Kidapawan Doctors College, IncDocument12 pagesKidapawan Doctors College, IncDonna AlejoNo ratings yet
- Unit 1 Anatomy, Physiology and The Human Body: Learning OutcomesDocument91 pagesUnit 1 Anatomy, Physiology and The Human Body: Learning OutcomesDeolita BadiangNo ratings yet
- Deuterostomia Wanninger2015Document220 pagesDeuterostomia Wanninger2015Isabella Maria Trujillo Pulgar100% (1)
- Anatomy and Physiology Nursing Mnemonics & TipsDocument5 pagesAnatomy and Physiology Nursing Mnemonics & TipsChris ZantiraNo ratings yet
- 38 - Epithelial Attachment and Downgrowth On Dental ImplantDocument9 pages38 - Epithelial Attachment and Downgrowth On Dental ImplantPablo BenitezNo ratings yet
- Estimation of Time Since Death Ba PDFDocument6 pagesEstimation of Time Since Death Ba PDFCesar Augusto Rojas MachucaNo ratings yet
- Advanced Drug Delivery Reviews Volume 64 IssueDocument10 pagesAdvanced Drug Delivery Reviews Volume 64 IssueNirvani HarisNo ratings yet
- John FerrisDocument485 pagesJohn Ferrissteptaker10No ratings yet
- Development of Tooth and Its Supporting TissuesDocument68 pagesDevelopment of Tooth and Its Supporting TissuesSaud IqbalNo ratings yet
- Test Bank For Human Anatomy 8th Edition by MartiniDocument25 pagesTest Bank For Human Anatomy 8th Edition by Martinisophietryphenaqnmms5No ratings yet
- Vesicular, Ulcerative, and Necrotic Dermatitis of ReptilesDocument21 pagesVesicular, Ulcerative, and Necrotic Dermatitis of ReptilesAlice RmaNo ratings yet
- Tyldesley's Oral MedicineDocument269 pagesTyldesley's Oral MedicinePierpaolo PaniNo ratings yet
- Epithelial TissueDocument5 pagesEpithelial TissueAliyah ZalzosNo ratings yet
- Animal Tissues 22Document8 pagesAnimal Tissues 22Arleen ColacionNo ratings yet
- IX ABC SCI Final QP 23-24Document5 pagesIX ABC SCI Final QP 23-24Brijesh MishraNo ratings yet
- 1) Cell Injury BPT - 092731Document110 pages1) Cell Injury BPT - 092731Crystal GamingNo ratings yet
- Chapter 15 Innate ImmunityDocument38 pagesChapter 15 Innate Immunityapi-270625839No ratings yet
- Important Short Questions & Answers: Human Anatomy & Physiology HAP 20114Document12 pagesImportant Short Questions & Answers: Human Anatomy & Physiology HAP 20114Kunal DangatNo ratings yet
- Cell Modifications That Lead To AdaptationDocument1 pageCell Modifications That Lead To AdaptationJohn Barry Ibanez80% (15)
- DR Agarwals Textbook On Contact LensesDocument312 pagesDR Agarwals Textbook On Contact LensesManish MahabirNo ratings yet
- Test Questions: A. Cardiovascular System and LymphaticsDocument24 pagesTest Questions: A. Cardiovascular System and LymphaticsLunna Cruz100% (1)
- BYJU'S Subjective Mock Paper DiscussionDocument7 pagesBYJU'S Subjective Mock Paper DiscussionNaman VatsNo ratings yet
- Animal Tissues - Micro Notes by Seep PahujaDocument3 pagesAnimal Tissues - Micro Notes by Seep Pahujahh100% (1)
- Urmium Eng - 1 - CTYDocument50 pagesUrmium Eng - 1 - CTYnguyenthequang03No ratings yet