
PER Report Eli Scharlat 341394757 Final (2)
PER Report Eli Scharlat 341394757 Final (2)
You might also like
- LOV-based Optogenetic ToolsDocument36 pagesLOV-based Optogenetic ToolsAdam ZHUNo ratings yet
- Analyzing DataDocument4 pagesAnalyzing DataCarlos Matute NeavesNo ratings yet
- DOE Mentos and Soda Final PaperDocument9 pagesDOE Mentos and Soda Final Paperjxl6599No ratings yet
- Upt Puskesmas Atap: Pemerintah Kabupaten Nunukan Dinas KesehatanDocument19 pagesUpt Puskesmas Atap: Pemerintah Kabupaten Nunukan Dinas KesehatanIin SulmawatiNo ratings yet
- Pigmentation and Delayed Oxidation of Broiler ChickensDocument6 pagesPigmentation and Delayed Oxidation of Broiler ChickensAndrea SallayNo ratings yet
- A28 (Tavuk Karaciğer Kemik EurPoulSci)Document9 pagesA28 (Tavuk Karaciğer Kemik EurPoulSci)cht aytNo ratings yet
- Workshop 2 PDFDocument2 pagesWorkshop 2 PDFJosieNo ratings yet
- Lab Report Enzyme KineticsDocument5 pagesLab Report Enzyme KineticsABEGAIL JOY MAURICIO LAPITANNo ratings yet
- Food Nutrient Analysis LabDocument2 pagesFood Nutrient Analysis LabHansa BorichaNo ratings yet
- Assessment of The Minimal Available Phosphorus Needs of Laying - 2018 - PoultryDocument11 pagesAssessment of The Minimal Available Phosphorus Needs of Laying - 2018 - Poultrylady mae rufinoNo ratings yet
- Departments of Dairy and Agricultural Chemistry, Michigan State Ltniversity, T Ast LansingDocument5 pagesDepartments of Dairy and Agricultural Chemistry, Michigan State Ltniversity, T Ast LansingKevin JunkarinNo ratings yet
- Effect of SnailDocument13 pagesEffect of SnailElvy ChardilaNo ratings yet
- PIIS0022030254912254Document4 pagesPIIS0022030254912254Banji MaikaNo ratings yet
- Study of Antimicrobial Activity From GuavaDocument9 pagesStudy of Antimicrobial Activity From GuavaAngelene NacarNo ratings yet
- Haematological and Serum Biochemical Indices of Starter Broilers Fed Leaf Meal of Neem (Azadirachta Indica)Document9 pagesHaematological and Serum Biochemical Indices of Starter Broilers Fed Leaf Meal of Neem (Azadirachta Indica)Eduard MorenoNo ratings yet
- 1998-Callus Induction and Somatic Embryogenesis of Phalaenopsis PDFDocument5 pages1998-Callus Induction and Somatic Embryogenesis of Phalaenopsis PDFRobinson TroncosNo ratings yet
- Serologic Test For Human Seminal FluidDocument6 pagesSerologic Test For Human Seminal FluidGleiza GasalaoNo ratings yet
- Perawatan BBL Mei 2021-1Document2 pagesPerawatan BBL Mei 2021-1wardaniNo ratings yet
- Tugas Devi SusantiDocument10 pagesTugas Devi SusantiNia Riski Aspriyanti linggauNo ratings yet
- Broiler SBA 2020-2021 A. Chambers - Brown-1Document14 pagesBroiler SBA 2020-2021 A. Chambers - Brown-1ChadNo ratings yet
- Bovine Virus Diarrhea (BVD) Dan MikoplasmaDocument6 pagesBovine Virus Diarrhea (BVD) Dan MikoplasmaRantika KhumairahNo ratings yet
- 10 Markah / 10 MarksDocument15 pages10 Markah / 10 MarksTc Nur 'AinNo ratings yet
- 1 s2.0 S0032579119388509 MainDocument15 pages1 s2.0 S0032579119388509 MainAlfred BaitanuNo ratings yet
- Lab 3Document4 pagesLab 3api-383615212No ratings yet
- Production Performance, Immune Response and Carcass Traits of Broiler Chickens Fed Diet Incorporated With Kappaphycus AlvareziiDocument8 pagesProduction Performance, Immune Response and Carcass Traits of Broiler Chickens Fed Diet Incorporated With Kappaphycus AlvareziiShiela Mae Gocotano BolambotNo ratings yet
- Biology Depth StudyDocument10 pagesBiology Depth StudyHira AliNo ratings yet
- Drosophilia Lab ReportDocument5 pagesDrosophilia Lab ReportMaddie BleaseNo ratings yet
- Social Defeat ParadigmDocument10 pagesSocial Defeat ParadigmShobhitNo ratings yet
- NSEJS 2023 Exam Paper SolutionDocument21 pagesNSEJS 2023 Exam Paper SolutionAaditya ThapaNo ratings yet
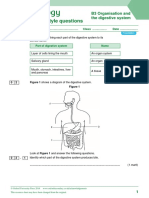
- Exam-Style Questions: B3 Organisation and The Digestive SystemDocument6 pagesExam-Style Questions: B3 Organisation and The Digestive Systemchemphycomb100% (1)
- Effects of Dietary Soybean Curd Residue On The Growth Performance and Carcass Characteristics in Hanwoo (Bos Taurus Coreanae) SteerDocument6 pagesEffects of Dietary Soybean Curd Residue On The Growth Performance and Carcass Characteristics in Hanwoo (Bos Taurus Coreanae) SteerGaryong VlogNo ratings yet
- To See OnDocument5 pagesTo See OnJames PerianayagamNo ratings yet
- Taqman Sample-To-Snp™ Kit: Prepare The SamplesDocument6 pagesTaqman Sample-To-Snp™ Kit: Prepare The Samplesjean pauloNo ratings yet
- Protein TechniquesDocument13 pagesProtein TechniquesRendel GuevarraNo ratings yet
- 1.beneficial Effects of Aloe Vera Leaf Gel Extract On Lipid Profile Status in Rats With Streptozotocin DiabetesDocument9 pages1.beneficial Effects of Aloe Vera Leaf Gel Extract On Lipid Profile Status in Rats With Streptozotocin DiabetesEric GibsonNo ratings yet
- Proprietary Content.: Great Learning. All Right Reserved. Unauthorized Use or Distribution ProhibitedDocument4 pagesProprietary Content.: Great Learning. All Right Reserved. Unauthorized Use or Distribution ProhibitedRishi Mukherjee0% (1)
- PharmacologyDocument11 pagesPharmacologylyracojaquilmacNo ratings yet
- Case 3-TuDocument4 pagesCase 3-TuGia HoàngNo ratings yet
- Bio Report (1) PrinttDocument5 pagesBio Report (1) PrinttNatalie OrtizNo ratings yet
- TD 4 CorrectionDocument6 pagesTD 4 Correctionahmedleo10010No ratings yet
- MPN Method BTS TXDocument7 pagesMPN Method BTS TXMuhammad Hanif GhifariNo ratings yet
- Effects of Different Feed Forms On Performance in Broiler ChickensDocument5 pagesEffects of Different Feed Forms On Performance in Broiler ChickensKhaeraniMahdinurAwliaNo ratings yet
- Pictures and List of Required Examples of Compliance Evidences ECE's in Physical Plant and Instructional Support Facilities 1. Building PlanDocument36 pagesPictures and List of Required Examples of Compliance Evidences ECE's in Physical Plant and Instructional Support Facilities 1. Building PlanEdel Mae Opena100% (1)
- Exercise 4: Data Summarizing and Presenting: JawabDocument10 pagesExercise 4: Data Summarizing and Presenting: Jawabwidya riliadhaNo ratings yet
- CHALLENGENaked PotatoesDocument3 pagesCHALLENGENaked PotatoesFernando NicastriNo ratings yet
- Experiment 3. The Test For Protein: Substance Reaction With Copper Sulphate and Sodium HydroxideDocument4 pagesExperiment 3. The Test For Protein: Substance Reaction With Copper Sulphate and Sodium HydroxideAndrew PooleNo ratings yet
- SPSS Tugas Nia Riski ADocument10 pagesSPSS Tugas Nia Riski ANia Riski Aspriyanti linggauNo ratings yet
- OK - Performance of Broilers Fed Premix From Locally Sourced MaterialsDocument4 pagesOK - Performance of Broilers Fed Premix From Locally Sourced MaterialsOliver TalipNo ratings yet
- Taxonomy: Bio 160 Unit 3 Study Guide VocabularyDocument11 pagesTaxonomy: Bio 160 Unit 3 Study Guide Vocabularyf7ktpjb6npNo ratings yet
- Lab 3 Standard Protein Assay LabDocument5 pagesLab 3 Standard Protein Assay LabMark WayneNo ratings yet
- 261 Labs - U1 - Osmolarity - Lab ReportDocument12 pages261 Labs - U1 - Osmolarity - Lab ReportSummerNo ratings yet
- Gel AgaroseDocument6 pagesGel Agarosemozhi74826207No ratings yet
- Finch Beak ActivityDocument3 pagesFinch Beak Activityapi-646571488No ratings yet
- El-Tahan-EFECT OF DIETARY STARCH LEVELS ON PERFORMANCE AND DIGESTIBILITY OF GROWING RABBITSDocument5 pagesEl-Tahan-EFECT OF DIETARY STARCH LEVELS ON PERFORMANCE AND DIGESTIBILITY OF GROWING RABBITSanayeli vazquez valladolidNo ratings yet
- Set 05 SolutionDocument5 pagesSet 05 Solutionnuravdoug8054No ratings yet
- Changes in Cecum Microbial Community in Response To Total Sulfur 2019 PoultrDocument11 pagesChanges in Cecum Microbial Community in Response To Total Sulfur 2019 PoultrAleneNo ratings yet
- Analisis Penilaian HarianDocument3 pagesAnalisis Penilaian HarianTrisnaYati Nona KarmadiNo ratings yet
- Tabulasi Terbaru Fix UdahDocument21 pagesTabulasi Terbaru Fix UdahNurika ListyanNo ratings yet
- Bio462 Practical 3Document10 pagesBio462 Practical 3kimnamin9No ratings yet
- A31 (Tavuk Yumurta Kalite Kafes Sis, AÜVF)Document5 pagesA31 (Tavuk Yumurta Kalite Kafes Sis, AÜVF)cht aytNo ratings yet
- The Fundamentals of Scientific Research: An Introductory Laboratory ManualFrom EverandThe Fundamentals of Scientific Research: An Introductory Laboratory ManualNo ratings yet
- Department of Education: Parental ConsentDocument10 pagesDepartment of Education: Parental ConsentESTER EVON MEDRANONo ratings yet
- Test Bank For Marriages Families and Relationships Making Choices in A Diverse Society 11 Edition Mary Ann LamannaDocument41 pagesTest Bank For Marriages Families and Relationships Making Choices in A Diverse Society 11 Edition Mary Ann Lamannaparchisidewg0drNo ratings yet
- Poly-L-Lysine Solution (0.01%) : Cell Culture Reagent Cat. # A-005-C Pack Size: 100mLDocument2 pagesPoly-L-Lysine Solution (0.01%) : Cell Culture Reagent Cat. # A-005-C Pack Size: 100mLConstanza NegreteNo ratings yet
- Bio 101 StemDocument52 pagesBio 101 StemLance CarandangNo ratings yet
- IAL Biology SB2 Answers 5BDocument6 pagesIAL Biology SB2 Answers 5BsalmaNo ratings yet
- Ben Hunt Grant Williams - 4Document5 pagesBen Hunt Grant Williams - 4Bob FisherNo ratings yet
- Nadia Handayani - 2213091007 - Artikel InggrisDocument4 pagesNadia Handayani - 2213091007 - Artikel InggrisNadia HandayaniNo ratings yet
- Tendon Transfers: by DR Krishna BhattDocument89 pagesTendon Transfers: by DR Krishna Bhattvenkata ramakrishnaiahNo ratings yet
- Embryonic DevelopmentDocument15 pagesEmbryonic DevelopmentАдия АбуNo ratings yet
- Research: Quarter 1 - Module 9 The Research ProblemDocument19 pagesResearch: Quarter 1 - Module 9 The Research ProblemKeziah LapojaNo ratings yet
- Group AssDocument21 pagesGroup AssAmanuel MergaNo ratings yet
- Tissues Level OrganizationDocument60 pagesTissues Level OrganizationHilman YusriNo ratings yet
- MMW The Nature of MathematicsDocument55 pagesMMW The Nature of MathematicsJessa Tangonan LauretaNo ratings yet
- Linkage and Genetic Mapping EukaryotesDocument37 pagesLinkage and Genetic Mapping EukaryotesSITI BAZILAH BINTI BILAK KPM-GuruNo ratings yet
- SSC-I Biology (2nd Set)Document11 pagesSSC-I Biology (2nd Set)Faisal SamiNo ratings yet
- TEST03Document2 pagesTEST03JoseMa AralNo ratings yet
- Activity No. 4: Simple and Complex TissuesDocument6 pagesActivity No. 4: Simple and Complex Tissuesmasor0% (1)
- Extraction and Characterization of Hyaluronic Acid From The Eyeball of Nile Tilapia Oreochromis Niloticus 2023Document12 pagesExtraction and Characterization of Hyaluronic Acid From The Eyeball of Nile Tilapia Oreochromis Niloticus 2023pablo.busattoNo ratings yet
- Lab Practical 3 - Manual FK10403 Wood Science - OnlineDocument22 pagesLab Practical 3 - Manual FK10403 Wood Science - OnlineYENNITA JAMES BF21110144No ratings yet
- Very Old Dinosaur-KeyDocument26 pagesVery Old Dinosaur-KeyedjlohouedanouNo ratings yet
- Full Ebook of Algorithms in Bioinformatics Theory and Implementation 1St Edition Gagniuc Paul A Online PDF All ChapterDocument69 pagesFull Ebook of Algorithms in Bioinformatics Theory and Implementation 1St Edition Gagniuc Paul A Online PDF All Chapterbaligiainiey100% (17)
- Go With Your Safety! GO IN-AC-TION!: Science 7Document15 pagesGo With Your Safety! GO IN-AC-TION!: Science 7Core GirlNo ratings yet
- Bio 30 Ol LRP 15 16Document4 pagesBio 30 Ol LRP 15 16danielNo ratings yet
- Evolution Part 1 by M.Saidi-coDocument23 pagesEvolution Part 1 by M.Saidi-coSIBONGILE FUNDENo ratings yet
- 345Document10 pages345Ariana ChimiNo ratings yet
- Act Sheet 15 Systematic Zoology 01 ExerciseDocument8 pagesAct Sheet 15 Systematic Zoology 01 ExerciseDusseldon ValenciaNo ratings yet
- Cell Structure and Function Multiple Choice Quiz Questions and AnswersDocument32 pagesCell Structure and Function Multiple Choice Quiz Questions and AnswersAlyssa Kim Fernandez50% (4)
- Human Germline Genome EditingDocument11 pagesHuman Germline Genome EditingSara Helena Cardona ToroNo ratings yet
- LRP 6 Q4 285 Pages in AllDocument96 pagesLRP 6 Q4 285 Pages in Allabracodera.bdcNo ratings yet

























































































Download as docx, pdf, or txt
You might also like
- LOV-based Optogenetic ToolsDocument36 pagesLOV-based Optogenetic ToolsAdam ZHUNo ratings yet
- Analyzing DataDocument4 pagesAnalyzing DataCarlos Matute NeavesNo ratings yet
- DOE Mentos and Soda Final PaperDocument9 pagesDOE Mentos and Soda Final Paperjxl6599No ratings yet
- Upt Puskesmas Atap: Pemerintah Kabupaten Nunukan Dinas KesehatanDocument19 pagesUpt Puskesmas Atap: Pemerintah Kabupaten Nunukan Dinas KesehatanIin SulmawatiNo ratings yet
- Pigmentation and Delayed Oxidation of Broiler ChickensDocument6 pagesPigmentation and Delayed Oxidation of Broiler ChickensAndrea SallayNo ratings yet
- A28 (Tavuk Karaciğer Kemik EurPoulSci)Document9 pagesA28 (Tavuk Karaciğer Kemik EurPoulSci)cht aytNo ratings yet
- Workshop 2 PDFDocument2 pagesWorkshop 2 PDFJosieNo ratings yet
- Lab Report Enzyme KineticsDocument5 pagesLab Report Enzyme KineticsABEGAIL JOY MAURICIO LAPITANNo ratings yet
- Food Nutrient Analysis LabDocument2 pagesFood Nutrient Analysis LabHansa BorichaNo ratings yet
- Assessment of The Minimal Available Phosphorus Needs of Laying - 2018 - PoultryDocument11 pagesAssessment of The Minimal Available Phosphorus Needs of Laying - 2018 - Poultrylady mae rufinoNo ratings yet
- Departments of Dairy and Agricultural Chemistry, Michigan State Ltniversity, T Ast LansingDocument5 pagesDepartments of Dairy and Agricultural Chemistry, Michigan State Ltniversity, T Ast LansingKevin JunkarinNo ratings yet
- Effect of SnailDocument13 pagesEffect of SnailElvy ChardilaNo ratings yet
- PIIS0022030254912254Document4 pagesPIIS0022030254912254Banji MaikaNo ratings yet
- Study of Antimicrobial Activity From GuavaDocument9 pagesStudy of Antimicrobial Activity From GuavaAngelene NacarNo ratings yet
- Haematological and Serum Biochemical Indices of Starter Broilers Fed Leaf Meal of Neem (Azadirachta Indica)Document9 pagesHaematological and Serum Biochemical Indices of Starter Broilers Fed Leaf Meal of Neem (Azadirachta Indica)Eduard MorenoNo ratings yet
- 1998-Callus Induction and Somatic Embryogenesis of Phalaenopsis PDFDocument5 pages1998-Callus Induction and Somatic Embryogenesis of Phalaenopsis PDFRobinson TroncosNo ratings yet
- Serologic Test For Human Seminal FluidDocument6 pagesSerologic Test For Human Seminal FluidGleiza GasalaoNo ratings yet
- Perawatan BBL Mei 2021-1Document2 pagesPerawatan BBL Mei 2021-1wardaniNo ratings yet
- Tugas Devi SusantiDocument10 pagesTugas Devi SusantiNia Riski Aspriyanti linggauNo ratings yet
- Broiler SBA 2020-2021 A. Chambers - Brown-1Document14 pagesBroiler SBA 2020-2021 A. Chambers - Brown-1ChadNo ratings yet
- Bovine Virus Diarrhea (BVD) Dan MikoplasmaDocument6 pagesBovine Virus Diarrhea (BVD) Dan MikoplasmaRantika KhumairahNo ratings yet
- 10 Markah / 10 MarksDocument15 pages10 Markah / 10 MarksTc Nur 'AinNo ratings yet
- 1 s2.0 S0032579119388509 MainDocument15 pages1 s2.0 S0032579119388509 MainAlfred BaitanuNo ratings yet
- Lab 3Document4 pagesLab 3api-383615212No ratings yet
- Production Performance, Immune Response and Carcass Traits of Broiler Chickens Fed Diet Incorporated With Kappaphycus AlvareziiDocument8 pagesProduction Performance, Immune Response and Carcass Traits of Broiler Chickens Fed Diet Incorporated With Kappaphycus AlvareziiShiela Mae Gocotano BolambotNo ratings yet
- Biology Depth StudyDocument10 pagesBiology Depth StudyHira AliNo ratings yet
- Drosophilia Lab ReportDocument5 pagesDrosophilia Lab ReportMaddie BleaseNo ratings yet
- Social Defeat ParadigmDocument10 pagesSocial Defeat ParadigmShobhitNo ratings yet
- NSEJS 2023 Exam Paper SolutionDocument21 pagesNSEJS 2023 Exam Paper SolutionAaditya ThapaNo ratings yet
- Exam-Style Questions: B3 Organisation and The Digestive SystemDocument6 pagesExam-Style Questions: B3 Organisation and The Digestive Systemchemphycomb100% (1)
- Effects of Dietary Soybean Curd Residue On The Growth Performance and Carcass Characteristics in Hanwoo (Bos Taurus Coreanae) SteerDocument6 pagesEffects of Dietary Soybean Curd Residue On The Growth Performance and Carcass Characteristics in Hanwoo (Bos Taurus Coreanae) SteerGaryong VlogNo ratings yet
- To See OnDocument5 pagesTo See OnJames PerianayagamNo ratings yet
- Taqman Sample-To-Snp™ Kit: Prepare The SamplesDocument6 pagesTaqman Sample-To-Snp™ Kit: Prepare The Samplesjean pauloNo ratings yet
- Protein TechniquesDocument13 pagesProtein TechniquesRendel GuevarraNo ratings yet
- 1.beneficial Effects of Aloe Vera Leaf Gel Extract On Lipid Profile Status in Rats With Streptozotocin DiabetesDocument9 pages1.beneficial Effects of Aloe Vera Leaf Gel Extract On Lipid Profile Status in Rats With Streptozotocin DiabetesEric GibsonNo ratings yet
- Proprietary Content.: Great Learning. All Right Reserved. Unauthorized Use or Distribution ProhibitedDocument4 pagesProprietary Content.: Great Learning. All Right Reserved. Unauthorized Use or Distribution ProhibitedRishi Mukherjee0% (1)
- PharmacologyDocument11 pagesPharmacologylyracojaquilmacNo ratings yet
- Case 3-TuDocument4 pagesCase 3-TuGia HoàngNo ratings yet
- Bio Report (1) PrinttDocument5 pagesBio Report (1) PrinttNatalie OrtizNo ratings yet
- TD 4 CorrectionDocument6 pagesTD 4 Correctionahmedleo10010No ratings yet
- MPN Method BTS TXDocument7 pagesMPN Method BTS TXMuhammad Hanif GhifariNo ratings yet
- Effects of Different Feed Forms On Performance in Broiler ChickensDocument5 pagesEffects of Different Feed Forms On Performance in Broiler ChickensKhaeraniMahdinurAwliaNo ratings yet
- Pictures and List of Required Examples of Compliance Evidences ECE's in Physical Plant and Instructional Support Facilities 1. Building PlanDocument36 pagesPictures and List of Required Examples of Compliance Evidences ECE's in Physical Plant and Instructional Support Facilities 1. Building PlanEdel Mae Opena100% (1)
- Exercise 4: Data Summarizing and Presenting: JawabDocument10 pagesExercise 4: Data Summarizing and Presenting: Jawabwidya riliadhaNo ratings yet
- CHALLENGENaked PotatoesDocument3 pagesCHALLENGENaked PotatoesFernando NicastriNo ratings yet
- Experiment 3. The Test For Protein: Substance Reaction With Copper Sulphate and Sodium HydroxideDocument4 pagesExperiment 3. The Test For Protein: Substance Reaction With Copper Sulphate and Sodium HydroxideAndrew PooleNo ratings yet
- SPSS Tugas Nia Riski ADocument10 pagesSPSS Tugas Nia Riski ANia Riski Aspriyanti linggauNo ratings yet
- OK - Performance of Broilers Fed Premix From Locally Sourced MaterialsDocument4 pagesOK - Performance of Broilers Fed Premix From Locally Sourced MaterialsOliver TalipNo ratings yet
- Taxonomy: Bio 160 Unit 3 Study Guide VocabularyDocument11 pagesTaxonomy: Bio 160 Unit 3 Study Guide Vocabularyf7ktpjb6npNo ratings yet
- Lab 3 Standard Protein Assay LabDocument5 pagesLab 3 Standard Protein Assay LabMark WayneNo ratings yet
- 261 Labs - U1 - Osmolarity - Lab ReportDocument12 pages261 Labs - U1 - Osmolarity - Lab ReportSummerNo ratings yet
- Gel AgaroseDocument6 pagesGel Agarosemozhi74826207No ratings yet
- Finch Beak ActivityDocument3 pagesFinch Beak Activityapi-646571488No ratings yet
- El-Tahan-EFECT OF DIETARY STARCH LEVELS ON PERFORMANCE AND DIGESTIBILITY OF GROWING RABBITSDocument5 pagesEl-Tahan-EFECT OF DIETARY STARCH LEVELS ON PERFORMANCE AND DIGESTIBILITY OF GROWING RABBITSanayeli vazquez valladolidNo ratings yet
- Set 05 SolutionDocument5 pagesSet 05 Solutionnuravdoug8054No ratings yet
- Changes in Cecum Microbial Community in Response To Total Sulfur 2019 PoultrDocument11 pagesChanges in Cecum Microbial Community in Response To Total Sulfur 2019 PoultrAleneNo ratings yet
- Analisis Penilaian HarianDocument3 pagesAnalisis Penilaian HarianTrisnaYati Nona KarmadiNo ratings yet
- Tabulasi Terbaru Fix UdahDocument21 pagesTabulasi Terbaru Fix UdahNurika ListyanNo ratings yet
- Bio462 Practical 3Document10 pagesBio462 Practical 3kimnamin9No ratings yet
- A31 (Tavuk Yumurta Kalite Kafes Sis, AÜVF)Document5 pagesA31 (Tavuk Yumurta Kalite Kafes Sis, AÜVF)cht aytNo ratings yet
- The Fundamentals of Scientific Research: An Introductory Laboratory ManualFrom EverandThe Fundamentals of Scientific Research: An Introductory Laboratory ManualNo ratings yet
- Department of Education: Parental ConsentDocument10 pagesDepartment of Education: Parental ConsentESTER EVON MEDRANONo ratings yet
- Test Bank For Marriages Families and Relationships Making Choices in A Diverse Society 11 Edition Mary Ann LamannaDocument41 pagesTest Bank For Marriages Families and Relationships Making Choices in A Diverse Society 11 Edition Mary Ann Lamannaparchisidewg0drNo ratings yet
- Poly-L-Lysine Solution (0.01%) : Cell Culture Reagent Cat. # A-005-C Pack Size: 100mLDocument2 pagesPoly-L-Lysine Solution (0.01%) : Cell Culture Reagent Cat. # A-005-C Pack Size: 100mLConstanza NegreteNo ratings yet
- Bio 101 StemDocument52 pagesBio 101 StemLance CarandangNo ratings yet
- IAL Biology SB2 Answers 5BDocument6 pagesIAL Biology SB2 Answers 5BsalmaNo ratings yet
- Ben Hunt Grant Williams - 4Document5 pagesBen Hunt Grant Williams - 4Bob FisherNo ratings yet
- Nadia Handayani - 2213091007 - Artikel InggrisDocument4 pagesNadia Handayani - 2213091007 - Artikel InggrisNadia HandayaniNo ratings yet
- Tendon Transfers: by DR Krishna BhattDocument89 pagesTendon Transfers: by DR Krishna Bhattvenkata ramakrishnaiahNo ratings yet
- Embryonic DevelopmentDocument15 pagesEmbryonic DevelopmentАдия АбуNo ratings yet
- Research: Quarter 1 - Module 9 The Research ProblemDocument19 pagesResearch: Quarter 1 - Module 9 The Research ProblemKeziah LapojaNo ratings yet
- Group AssDocument21 pagesGroup AssAmanuel MergaNo ratings yet
- Tissues Level OrganizationDocument60 pagesTissues Level OrganizationHilman YusriNo ratings yet
- MMW The Nature of MathematicsDocument55 pagesMMW The Nature of MathematicsJessa Tangonan LauretaNo ratings yet
- Linkage and Genetic Mapping EukaryotesDocument37 pagesLinkage and Genetic Mapping EukaryotesSITI BAZILAH BINTI BILAK KPM-GuruNo ratings yet
- SSC-I Biology (2nd Set)Document11 pagesSSC-I Biology (2nd Set)Faisal SamiNo ratings yet
- TEST03Document2 pagesTEST03JoseMa AralNo ratings yet
- Activity No. 4: Simple and Complex TissuesDocument6 pagesActivity No. 4: Simple and Complex Tissuesmasor0% (1)
- Extraction and Characterization of Hyaluronic Acid From The Eyeball of Nile Tilapia Oreochromis Niloticus 2023Document12 pagesExtraction and Characterization of Hyaluronic Acid From The Eyeball of Nile Tilapia Oreochromis Niloticus 2023pablo.busattoNo ratings yet
- Lab Practical 3 - Manual FK10403 Wood Science - OnlineDocument22 pagesLab Practical 3 - Manual FK10403 Wood Science - OnlineYENNITA JAMES BF21110144No ratings yet
- Very Old Dinosaur-KeyDocument26 pagesVery Old Dinosaur-KeyedjlohouedanouNo ratings yet
- Full Ebook of Algorithms in Bioinformatics Theory and Implementation 1St Edition Gagniuc Paul A Online PDF All ChapterDocument69 pagesFull Ebook of Algorithms in Bioinformatics Theory and Implementation 1St Edition Gagniuc Paul A Online PDF All Chapterbaligiainiey100% (17)
- Go With Your Safety! GO IN-AC-TION!: Science 7Document15 pagesGo With Your Safety! GO IN-AC-TION!: Science 7Core GirlNo ratings yet
- Bio 30 Ol LRP 15 16Document4 pagesBio 30 Ol LRP 15 16danielNo ratings yet
- Evolution Part 1 by M.Saidi-coDocument23 pagesEvolution Part 1 by M.Saidi-coSIBONGILE FUNDENo ratings yet
- 345Document10 pages345Ariana ChimiNo ratings yet
- Act Sheet 15 Systematic Zoology 01 ExerciseDocument8 pagesAct Sheet 15 Systematic Zoology 01 ExerciseDusseldon ValenciaNo ratings yet
- Cell Structure and Function Multiple Choice Quiz Questions and AnswersDocument32 pagesCell Structure and Function Multiple Choice Quiz Questions and AnswersAlyssa Kim Fernandez50% (4)
- Human Germline Genome EditingDocument11 pagesHuman Germline Genome EditingSara Helena Cardona ToroNo ratings yet
- LRP 6 Q4 285 Pages in AllDocument96 pagesLRP 6 Q4 285 Pages in Allabracodera.bdcNo ratings yet