
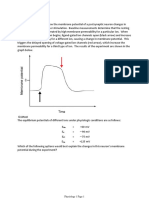
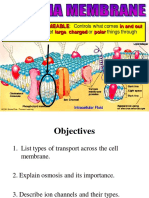
Chapter-8 enzymes
Chapter-8 enzymes
You might also like
- Cognitive Psychology and Its Implications 8th Edition Anderson Test BankDocument21 pagesCognitive Psychology and Its Implications 8th Edition Anderson Test Bankalexispatrickespgjiyntd100% (38)
- Unit 1 Bio Potential Generation and Electrode TypesDocument13 pagesUnit 1 Bio Potential Generation and Electrode TypesAleeshaNo ratings yet
- Introduction To RMP ND APDocument78 pagesIntroduction To RMP ND APHariharanNo ratings yet
- What Is Resting Membrane Potential?: Mechanism of An Action PotentialDocument5 pagesWhat Is Resting Membrane Potential?: Mechanism of An Action PotentialAli InamNo ratings yet
- 16.2 How Neurons Communicate - Concepts of BiologDocument5 pages16.2 How Neurons Communicate - Concepts of BiologjkhdgmbrvdNo ratings yet
- Electrical Properties of BiomembranesDocument52 pagesElectrical Properties of Biomembranesc3rberussNo ratings yet
- Passive Membrane PropertiesDocument10 pagesPassive Membrane PropertiesAhmed FrahatNo ratings yet
- Topic 3-Active Transport - EditedDocument7 pagesTopic 3-Active Transport - Editedsalve joy villanuevaNo ratings yet
- Excitable Tissue PhysiologyDocument13 pagesExcitable Tissue PhysiologyAlbert Che33% (3)
- 12 17 PDFDocument71 pages12 17 PDFVikiNo ratings yet
- Chapter 2 - Membrane PhysiologyDocument11 pagesChapter 2 - Membrane Physiologytahira UroojNo ratings yet
- Resting Membrane Potential Review QuestionsDocument3 pagesResting Membrane Potential Review QuestionsquyzbukNo ratings yet
- Types of Neuron Transmission and Communication SystemDocument5 pagesTypes of Neuron Transmission and Communication SystemRbbani AwanNo ratings yet
- Physiology Pearls by DR - SaqibDocument61 pagesPhysiology Pearls by DR - SaqibDr. Sarthak MishraNo ratings yet
- Questions ExplanationDocument17 pagesQuestions ExplanationnomintmNo ratings yet
- Key Points RevisionDocument124 pagesKey Points RevisionSumeyyah KemalNo ratings yet
- Action PotentialDocument4 pagesAction PotentialfarahNo ratings yet
- Basics of Biomedical Instrumentation NotesDocument24 pagesBasics of Biomedical Instrumentation NotesSteni GNo ratings yet
- Active TransportDocument10 pagesActive TransportSylvia DuzonNo ratings yet
- Sic 1311Document70 pagesSic 1311Shashi RamNo ratings yet
- Bio PsycheDocument16 pagesBio PsycheShruti DharNo ratings yet
- NEURONDocument4 pagesNEURONsummerbombabeNo ratings yet
- Neural TransmissionDocument21 pagesNeural TransmissionKawish AliNo ratings yet
- 7.7 - Membrane Potential - Chemistry LibreTextsDocument5 pages7.7 - Membrane Potential - Chemistry LibreTextswihiw92934No ratings yet
- Sodium Potassium Pump and Action PotentialDocument4 pagesSodium Potassium Pump and Action PotentialPiyush BhallaNo ratings yet
- Nerve and Muscle PhysiologyDocument82 pagesNerve and Muscle PhysiologychandsriNo ratings yet
- Action PotentialDocument2 pagesAction PotentialMian Talha KamalNo ratings yet
- Neuro 4Document11 pagesNeuro 4Marielle ZougheibNo ratings yet
- 1.0 The Origin of Biopotentials (New)Document62 pages1.0 The Origin of Biopotentials (New)Hanif HussinNo ratings yet
- Action Potential: L - T - P - S 3 - 0 - 2 - 0Document12 pagesAction Potential: L - T - P - S 3 - 0 - 2 - 0Ram MNo ratings yet
- The Action of Nerves: Appendix 4Document4 pagesThe Action of Nerves: Appendix 4natjeros12No ratings yet
- 2305 NeuronDocument5 pages2305 NeuronsridharNo ratings yet
- Action Potential: Nams, Bir Hospital B.SC - MIT 1 Year Suraj Sah Roll No. 9Document42 pagesAction Potential: Nams, Bir Hospital B.SC - MIT 1 Year Suraj Sah Roll No. 9Maxamed Cali AtomNo ratings yet
- 2305 NeuronDocument5 pages2305 NeuronAnwar AsyhraffNo ratings yet
- الوحدة الأولىDocument16 pagesالوحدة الأولىHUAWEI HUAWEINo ratings yet
- BIOC 315 - Lecture - 4Document29 pagesBIOC 315 - Lecture - 4Kevin KipropNo ratings yet
- Action Potential - The Resting Membrane Potential - Generation of Action Potentials - TeachMePhysiologAyDocument4 pagesAction Potential - The Resting Membrane Potential - Generation of Action Potentials - TeachMePhysiologAymohammed awolNo ratings yet
- Surgical PhsyiologyDocument170 pagesSurgical PhsyiologyTamjid Khan100% (2)
- Active TransportDocument51 pagesActive Transportericka khimNo ratings yet
- Nervous SystemDocument6 pagesNervous SystemarellanokristelleNo ratings yet
- Resting Membrane PotentialDocument8 pagesResting Membrane PotentialAhmad Atiq MalikiNo ratings yet
- RestingMembranePotential JeffLindoDocument8 pagesRestingMembranePotential JeffLindoClive NyabutiNo ratings yet
- Biopsych Notes p2Document28 pagesBiopsych Notes p2noorNo ratings yet
- RestingMembranePotential JeffLindoDocument8 pagesRestingMembranePotential JeffLindoAbraham AberaNo ratings yet
- Autonomic Nervous SystemDocument19 pagesAutonomic Nervous SystemAnuvabNo ratings yet
- Bpk105 Assignment 1Document2 pagesBpk105 Assignment 1DomstrongNo ratings yet
- Group 4B Millares Mapari Alisbo Simulating NeuronsDocument11 pagesGroup 4B Millares Mapari Alisbo Simulating NeuronsSean Nathan MapariNo ratings yet
- Nervous System Topic 2 Lecture 4 5Document22 pagesNervous System Topic 2 Lecture 4 5amjad.46987No ratings yet
- RestingMembranePotential RevisionDocument5 pagesRestingMembranePotential RevisionshaNo ratings yet
- Surgical PhsyiologyDocument170 pagesSurgical PhsyiologySteve FerisNo ratings yet
- Basics of Biomedical Instrumentation OMD551Document22 pagesBasics of Biomedical Instrumentation OMD551Sri ArunaaNo ratings yet
- Ch12 2018Document83 pagesCh12 2018김승윤No ratings yet
- Unit1 BERS Notes-Part2Document11 pagesUnit1 BERS Notes-Part2Mr. PavanNo ratings yet
- Negative Negative: Chapter Two: Electrical Signals in Nerve CellsDocument6 pagesNegative Negative: Chapter Two: Electrical Signals in Nerve CellsStacy BrenesNo ratings yet
- 2222 TransportDocument42 pages2222 TransportaddwrNo ratings yet
- Lecture 1Document14 pagesLecture 1Mahreen NoorNo ratings yet
- Neurons Resting PotentialDocument5 pagesNeurons Resting PotentialEstelle HagenNo ratings yet
- Introduction To NeurophysiologyDocument13 pagesIntroduction To Neurophysiologyvenkata ramakrishnaiahNo ratings yet
- Lesson 1: The History of Bioelectricity: Fundamentals of Neuroscience, Part 1: The Electrical Properties of The NeuronDocument4 pagesLesson 1: The History of Bioelectricity: Fundamentals of Neuroscience, Part 1: The Electrical Properties of The NeuronXyrille SantosNo ratings yet
- PhysioEx Exercise 3 Activity 4Document5 pagesPhysioEx Exercise 3 Activity 4edvin merida pro100% (1)
- 2016 Brain Bee Competitions Questions - DoDocument10 pages2016 Brain Bee Competitions Questions - DoughNo ratings yet
- Full Test Bank For Essentials of Psychology 7Th Edition Douglas A Bernstein PDF Docx Full Chapter ChapterDocument36 pagesFull Test Bank For Essentials of Psychology 7Th Edition Douglas A Bernstein PDF Docx Full Chapter Chapterbethmcneilysgbzqkpnw100% (16)
- Terminologies of ANNDocument3 pagesTerminologies of ANNrioraj0012No ratings yet
- BiophyDocument51 pagesBiophyAshleigh RodriguezNo ratings yet
- Biopsychology-Reviewer PDFDocument12 pagesBiopsychology-Reviewer PDFsherlynmaytiglaoNo ratings yet
- Synaptic Organization and PhysiologyDocument5 pagesSynaptic Organization and PhysiologyMohammad AliNo ratings yet
- The Autonomic Nervous System OverviewDocument39 pagesThe Autonomic Nervous System OverviewAhaisibwe GordonNo ratings yet
- Ch03 Questions Answer KeyDocument18 pagesCh03 Questions Answer KeyMichaelNo ratings yet
- Biological Psychology 11th Edition Kalat Test BankDocument25 pagesBiological Psychology 11th Edition Kalat Test BankMelissaJacksonDDSgwjp100% (63)
- Central and Peripheral Nervous SystemDocument36 pagesCentral and Peripheral Nervous SystemSehasa PathiranaNo ratings yet
- C 2.2 HL Neural SignalingDocument37 pagesC 2.2 HL Neural Signalingelizabeth.arockiadassNo ratings yet
- Lecture - Amygdala and Its FunctionsDocument8 pagesLecture - Amygdala and Its Functionsproblemchik2003100% (1)
- Memory Learning TechniqueDocument2 pagesMemory Learning Techniquerla97623No ratings yet
- Branches of BiopsychologyDocument4 pagesBranches of BiopsychologyPoulomiNo ratings yet
- Neurostimulation For Functional Recovery After.2Document8 pagesNeurostimulation For Functional Recovery After.2Mateo MendozaNo ratings yet
- Final Report BioDocument27 pagesFinal Report BioR FaNo ratings yet
- Unit 3Document62 pagesUnit 3Rishika ShrivastavaNo ratings yet
- Contemporary Psychiatric-Mental Health Nursing 3rd Edition Kneisl Solutions Manual DownloadDocument9 pagesContemporary Psychiatric-Mental Health Nursing 3rd Edition Kneisl Solutions Manual DownloadClarence Steele100% (24)
- Brain Computations and Connectivity Edmund T Rolls Full ChapterDocument51 pagesBrain Computations and Connectivity Edmund T Rolls Full Chaptertammy.wyatt492100% (6)
- Takakusaki Et Al 2022 - Neurophysiological Mechanisms of Gait Disturbance in Advanced ParkinsonDocument17 pagesTakakusaki Et Al 2022 - Neurophysiological Mechanisms of Gait Disturbance in Advanced ParkinsonlicmarinamaranNo ratings yet
- Functions and Properties of Synapse, ReflexDocument28 pagesFunctions and Properties of Synapse, ReflexAnupam MittalNo ratings yet
- Somatosensory CortexDocument19 pagesSomatosensory Cortexusamabangash943No ratings yet
- Gender Differences in Empathy: The Role of The Right Hemisphere: Rueckert Naybar 2008Document6 pagesGender Differences in Empathy: The Role of The Right Hemisphere: Rueckert Naybar 2008Morgan BenterudNo ratings yet
- Kami Export - 4) PHED 2506 - Lab 2 - Experiments WorksheetDocument12 pagesKami Export - 4) PHED 2506 - Lab 2 - Experiments WorksheetplerouxNo ratings yet
- Essentials of Understanding Psychology Canadian 5th Edition Feldman Test BankDocument30 pagesEssentials of Understanding Psychology Canadian 5th Edition Feldman Test BankEricFuentesfxgir100% (13)
- PerceptionDocument21 pagesPerceptionJaanu JaanuNo ratings yet
- Biological Basis of BehaviorDocument27 pagesBiological Basis of Behaviorhafizamanzoor44No ratings yet
- Practice Stimulus Response Model-TemplateDocument2 pagesPractice Stimulus Response Model-TemplateJohnny NguyenNo ratings yet
























































































Download as docx, pdf, or txt
You might also like
- Cognitive Psychology and Its Implications 8th Edition Anderson Test BankDocument21 pagesCognitive Psychology and Its Implications 8th Edition Anderson Test Bankalexispatrickespgjiyntd100% (38)
- Unit 1 Bio Potential Generation and Electrode TypesDocument13 pagesUnit 1 Bio Potential Generation and Electrode TypesAleeshaNo ratings yet
- Introduction To RMP ND APDocument78 pagesIntroduction To RMP ND APHariharanNo ratings yet
- What Is Resting Membrane Potential?: Mechanism of An Action PotentialDocument5 pagesWhat Is Resting Membrane Potential?: Mechanism of An Action PotentialAli InamNo ratings yet
- 16.2 How Neurons Communicate - Concepts of BiologDocument5 pages16.2 How Neurons Communicate - Concepts of BiologjkhdgmbrvdNo ratings yet
- Electrical Properties of BiomembranesDocument52 pagesElectrical Properties of Biomembranesc3rberussNo ratings yet
- Passive Membrane PropertiesDocument10 pagesPassive Membrane PropertiesAhmed FrahatNo ratings yet
- Topic 3-Active Transport - EditedDocument7 pagesTopic 3-Active Transport - Editedsalve joy villanuevaNo ratings yet
- Excitable Tissue PhysiologyDocument13 pagesExcitable Tissue PhysiologyAlbert Che33% (3)
- 12 17 PDFDocument71 pages12 17 PDFVikiNo ratings yet
- Chapter 2 - Membrane PhysiologyDocument11 pagesChapter 2 - Membrane Physiologytahira UroojNo ratings yet
- Resting Membrane Potential Review QuestionsDocument3 pagesResting Membrane Potential Review QuestionsquyzbukNo ratings yet
- Types of Neuron Transmission and Communication SystemDocument5 pagesTypes of Neuron Transmission and Communication SystemRbbani AwanNo ratings yet
- Physiology Pearls by DR - SaqibDocument61 pagesPhysiology Pearls by DR - SaqibDr. Sarthak MishraNo ratings yet
- Questions ExplanationDocument17 pagesQuestions ExplanationnomintmNo ratings yet
- Key Points RevisionDocument124 pagesKey Points RevisionSumeyyah KemalNo ratings yet
- Action PotentialDocument4 pagesAction PotentialfarahNo ratings yet
- Basics of Biomedical Instrumentation NotesDocument24 pagesBasics of Biomedical Instrumentation NotesSteni GNo ratings yet
- Active TransportDocument10 pagesActive TransportSylvia DuzonNo ratings yet
- Sic 1311Document70 pagesSic 1311Shashi RamNo ratings yet
- Bio PsycheDocument16 pagesBio PsycheShruti DharNo ratings yet
- NEURONDocument4 pagesNEURONsummerbombabeNo ratings yet
- Neural TransmissionDocument21 pagesNeural TransmissionKawish AliNo ratings yet
- 7.7 - Membrane Potential - Chemistry LibreTextsDocument5 pages7.7 - Membrane Potential - Chemistry LibreTextswihiw92934No ratings yet
- Sodium Potassium Pump and Action PotentialDocument4 pagesSodium Potassium Pump and Action PotentialPiyush BhallaNo ratings yet
- Nerve and Muscle PhysiologyDocument82 pagesNerve and Muscle PhysiologychandsriNo ratings yet
- Action PotentialDocument2 pagesAction PotentialMian Talha KamalNo ratings yet
- Neuro 4Document11 pagesNeuro 4Marielle ZougheibNo ratings yet
- 1.0 The Origin of Biopotentials (New)Document62 pages1.0 The Origin of Biopotentials (New)Hanif HussinNo ratings yet
- Action Potential: L - T - P - S 3 - 0 - 2 - 0Document12 pagesAction Potential: L - T - P - S 3 - 0 - 2 - 0Ram MNo ratings yet
- The Action of Nerves: Appendix 4Document4 pagesThe Action of Nerves: Appendix 4natjeros12No ratings yet
- 2305 NeuronDocument5 pages2305 NeuronsridharNo ratings yet
- Action Potential: Nams, Bir Hospital B.SC - MIT 1 Year Suraj Sah Roll No. 9Document42 pagesAction Potential: Nams, Bir Hospital B.SC - MIT 1 Year Suraj Sah Roll No. 9Maxamed Cali AtomNo ratings yet
- 2305 NeuronDocument5 pages2305 NeuronAnwar AsyhraffNo ratings yet
- الوحدة الأولىDocument16 pagesالوحدة الأولىHUAWEI HUAWEINo ratings yet
- BIOC 315 - Lecture - 4Document29 pagesBIOC 315 - Lecture - 4Kevin KipropNo ratings yet
- Action Potential - The Resting Membrane Potential - Generation of Action Potentials - TeachMePhysiologAyDocument4 pagesAction Potential - The Resting Membrane Potential - Generation of Action Potentials - TeachMePhysiologAymohammed awolNo ratings yet
- Surgical PhsyiologyDocument170 pagesSurgical PhsyiologyTamjid Khan100% (2)
- Active TransportDocument51 pagesActive Transportericka khimNo ratings yet
- Nervous SystemDocument6 pagesNervous SystemarellanokristelleNo ratings yet
- Resting Membrane PotentialDocument8 pagesResting Membrane PotentialAhmad Atiq MalikiNo ratings yet
- RestingMembranePotential JeffLindoDocument8 pagesRestingMembranePotential JeffLindoClive NyabutiNo ratings yet
- Biopsych Notes p2Document28 pagesBiopsych Notes p2noorNo ratings yet
- RestingMembranePotential JeffLindoDocument8 pagesRestingMembranePotential JeffLindoAbraham AberaNo ratings yet
- Autonomic Nervous SystemDocument19 pagesAutonomic Nervous SystemAnuvabNo ratings yet
- Bpk105 Assignment 1Document2 pagesBpk105 Assignment 1DomstrongNo ratings yet
- Group 4B Millares Mapari Alisbo Simulating NeuronsDocument11 pagesGroup 4B Millares Mapari Alisbo Simulating NeuronsSean Nathan MapariNo ratings yet
- Nervous System Topic 2 Lecture 4 5Document22 pagesNervous System Topic 2 Lecture 4 5amjad.46987No ratings yet
- RestingMembranePotential RevisionDocument5 pagesRestingMembranePotential RevisionshaNo ratings yet
- Surgical PhsyiologyDocument170 pagesSurgical PhsyiologySteve FerisNo ratings yet
- Basics of Biomedical Instrumentation OMD551Document22 pagesBasics of Biomedical Instrumentation OMD551Sri ArunaaNo ratings yet
- Ch12 2018Document83 pagesCh12 2018김승윤No ratings yet
- Unit1 BERS Notes-Part2Document11 pagesUnit1 BERS Notes-Part2Mr. PavanNo ratings yet
- Negative Negative: Chapter Two: Electrical Signals in Nerve CellsDocument6 pagesNegative Negative: Chapter Two: Electrical Signals in Nerve CellsStacy BrenesNo ratings yet
- 2222 TransportDocument42 pages2222 TransportaddwrNo ratings yet
- Lecture 1Document14 pagesLecture 1Mahreen NoorNo ratings yet
- Neurons Resting PotentialDocument5 pagesNeurons Resting PotentialEstelle HagenNo ratings yet
- Introduction To NeurophysiologyDocument13 pagesIntroduction To Neurophysiologyvenkata ramakrishnaiahNo ratings yet
- Lesson 1: The History of Bioelectricity: Fundamentals of Neuroscience, Part 1: The Electrical Properties of The NeuronDocument4 pagesLesson 1: The History of Bioelectricity: Fundamentals of Neuroscience, Part 1: The Electrical Properties of The NeuronXyrille SantosNo ratings yet
- PhysioEx Exercise 3 Activity 4Document5 pagesPhysioEx Exercise 3 Activity 4edvin merida pro100% (1)
- 2016 Brain Bee Competitions Questions - DoDocument10 pages2016 Brain Bee Competitions Questions - DoughNo ratings yet
- Full Test Bank For Essentials of Psychology 7Th Edition Douglas A Bernstein PDF Docx Full Chapter ChapterDocument36 pagesFull Test Bank For Essentials of Psychology 7Th Edition Douglas A Bernstein PDF Docx Full Chapter Chapterbethmcneilysgbzqkpnw100% (16)
- Terminologies of ANNDocument3 pagesTerminologies of ANNrioraj0012No ratings yet
- BiophyDocument51 pagesBiophyAshleigh RodriguezNo ratings yet
- Biopsychology-Reviewer PDFDocument12 pagesBiopsychology-Reviewer PDFsherlynmaytiglaoNo ratings yet
- Synaptic Organization and PhysiologyDocument5 pagesSynaptic Organization and PhysiologyMohammad AliNo ratings yet
- The Autonomic Nervous System OverviewDocument39 pagesThe Autonomic Nervous System OverviewAhaisibwe GordonNo ratings yet
- Ch03 Questions Answer KeyDocument18 pagesCh03 Questions Answer KeyMichaelNo ratings yet
- Biological Psychology 11th Edition Kalat Test BankDocument25 pagesBiological Psychology 11th Edition Kalat Test BankMelissaJacksonDDSgwjp100% (63)
- Central and Peripheral Nervous SystemDocument36 pagesCentral and Peripheral Nervous SystemSehasa PathiranaNo ratings yet
- C 2.2 HL Neural SignalingDocument37 pagesC 2.2 HL Neural Signalingelizabeth.arockiadassNo ratings yet
- Lecture - Amygdala and Its FunctionsDocument8 pagesLecture - Amygdala and Its Functionsproblemchik2003100% (1)
- Memory Learning TechniqueDocument2 pagesMemory Learning Techniquerla97623No ratings yet
- Branches of BiopsychologyDocument4 pagesBranches of BiopsychologyPoulomiNo ratings yet
- Neurostimulation For Functional Recovery After.2Document8 pagesNeurostimulation For Functional Recovery After.2Mateo MendozaNo ratings yet
- Final Report BioDocument27 pagesFinal Report BioR FaNo ratings yet
- Unit 3Document62 pagesUnit 3Rishika ShrivastavaNo ratings yet
- Contemporary Psychiatric-Mental Health Nursing 3rd Edition Kneisl Solutions Manual DownloadDocument9 pagesContemporary Psychiatric-Mental Health Nursing 3rd Edition Kneisl Solutions Manual DownloadClarence Steele100% (24)
- Brain Computations and Connectivity Edmund T Rolls Full ChapterDocument51 pagesBrain Computations and Connectivity Edmund T Rolls Full Chaptertammy.wyatt492100% (6)
- Takakusaki Et Al 2022 - Neurophysiological Mechanisms of Gait Disturbance in Advanced ParkinsonDocument17 pagesTakakusaki Et Al 2022 - Neurophysiological Mechanisms of Gait Disturbance in Advanced ParkinsonlicmarinamaranNo ratings yet
- Functions and Properties of Synapse, ReflexDocument28 pagesFunctions and Properties of Synapse, ReflexAnupam MittalNo ratings yet
- Somatosensory CortexDocument19 pagesSomatosensory Cortexusamabangash943No ratings yet
- Gender Differences in Empathy: The Role of The Right Hemisphere: Rueckert Naybar 2008Document6 pagesGender Differences in Empathy: The Role of The Right Hemisphere: Rueckert Naybar 2008Morgan BenterudNo ratings yet
- Kami Export - 4) PHED 2506 - Lab 2 - Experiments WorksheetDocument12 pagesKami Export - 4) PHED 2506 - Lab 2 - Experiments WorksheetplerouxNo ratings yet
- Essentials of Understanding Psychology Canadian 5th Edition Feldman Test BankDocument30 pagesEssentials of Understanding Psychology Canadian 5th Edition Feldman Test BankEricFuentesfxgir100% (13)
- PerceptionDocument21 pagesPerceptionJaanu JaanuNo ratings yet
- Biological Basis of BehaviorDocument27 pagesBiological Basis of Behaviorhafizamanzoor44No ratings yet
- Practice Stimulus Response Model-TemplateDocument2 pagesPractice Stimulus Response Model-TemplateJohnny NguyenNo ratings yet