Download as pdf or txt
You might also like
- Dihybrid ProblemsDocument7 pagesDihybrid Problemsbrenden chapmanNo ratings yet
- Injerto Tomate en Solanaceas - 2Document10 pagesInjerto Tomate en Solanaceas - 2Jeisson Daniel Torres QuinteroNo ratings yet
- 1983 2125 Rcaat 30 02 00391 PDFDocument10 pages1983 2125 Rcaat 30 02 00391 PDFElah DumlaoNo ratings yet
- tmp1F68 TMPDocument15 pagestmp1F68 TMPFrontiersNo ratings yet
- DownloadDocument11 pagesDownloadsgarbossa.jaquelineNo ratings yet
- Productivity and Quality of Formosa and SoloDocument8 pagesProductivity and Quality of Formosa and SoloJoão Carlos PinheiroNo ratings yet
- Physiology_development_in_the_vegetative_stage_of_Document11 pagesPhysiology_development_in_the_vegetative_stage_of_Risma Neswati SukimanNo ratings yet
- Cultivars Performance and Blackberry Pruning Management Desempeño de Cultivares y Manejo de Podas en MoraDocument15 pagesCultivars Performance and Blackberry Pruning Management Desempeño de Cultivares y Manejo de Podas en MoraSebastián BallénNo ratings yet
- Phytophthora Infestans Solanum LycopersiconDocument5 pagesPhytophthora Infestans Solanum LycopersiconHebert Jhimsy Pastor GarcíaNo ratings yet
- Artículo de Investigación: Laura Victoria PEREZ MARTINEZ, Luz Marina MELGAREJODocument12 pagesArtículo de Investigación: Laura Victoria PEREZ MARTINEZ, Luz Marina MELGAREJOEduardo DucuaraNo ratings yet
- Dosis Cafe CanephoraDocument8 pagesDosis Cafe CanephoraMelany NarváezNo ratings yet
- Melo Et Al., 2018 Feijão A&aDocument8 pagesMelo Et Al., 2018 Feijão A&aAlexson DutraNo ratings yet
- Mechanisms of Tolerance To Water Deficit and Physiological Responses To Rehydration in CowpeaDocument10 pagesMechanisms of Tolerance To Water Deficit and Physiological Responses To Rehydration in CowpeafranciscoNo ratings yet
- Castoldi 8 6 2014 901 908Document9 pagesCastoldi 8 6 2014 901 908JIMMY GERALD JUAREZ PAUCARNo ratings yet
- Carini Et AlDocument12 pagesCarini Et AlVirgilio Gavicho UarrotaNo ratings yet
- Acta Scientiarum 1Document11 pagesActa Scientiarum 1Manelli77No ratings yet
- Characterization of Leaves and Fruits of MangoDocument7 pagesCharacterization of Leaves and Fruits of MangoDaniel BartoloNo ratings yet
- SELECTION OF COFFEE PROGÊNIES FOR RESISTANCE TO LEAF RUDSiTas, A RN. DA. Et Al. FAVORABLE AGRONOMIC TRAITSDocument10 pagesSELECTION OF COFFEE PROGÊNIES FOR RESISTANCE TO LEAF RUDSiTas, A RN. DA. Et Al. FAVORABLE AGRONOMIC TRAITSMarceloNo ratings yet
- 2009 - Artigo 6- Physiological Analysis of Leaf SenescenceDocument6 pages2009 - Artigo 6- Physiological Analysis of Leaf Senescencevictoralyson360No ratings yet
- Theobroma Cacao Belongs To The Malvaceae Family and Subfamily Sterculioidea. It IsDocument18 pagesTheobroma Cacao Belongs To The Malvaceae Family and Subfamily Sterculioidea. It Isrendez vousNo ratings yet
- Maturação GuanandiDocument9 pagesMaturação GuanandiEvanny ValleriaNo ratings yet
- Effects of Pinching Time On The Performance of Three Tomato Varieties (Lycopersicon Lycopersicum Mill) in Ogbomoso and Mokwa, Nigeria'Document10 pagesEffects of Pinching Time On The Performance of Three Tomato Varieties (Lycopersicon Lycopersicum Mill) in Ogbomoso and Mokwa, Nigeria'IJEAB JournalNo ratings yet
- Spa No 2003Document6 pagesSpa No 2003uttam_bhuNo ratings yet
- Cercosporiose Progression in The Agroforestry Consortium Coffee-Rubber Trees Progresso Da Cercosporiose em Consórcio Agroflorestal Café-SeringueiraDocument10 pagesCercosporiose Progression in The Agroforestry Consortium Coffee-Rubber Trees Progresso Da Cercosporiose em Consórcio Agroflorestal Café-SeringueiraAzhari RizalNo ratings yet
- Initial Development and Tolerance of Pepper Species To Salinity StressDocument8 pagesInitial Development and Tolerance of Pepper Species To Salinity StressNiege FelixNo ratings yet
- Fisiologi Benih KopiDocument25 pagesFisiologi Benih KopiAlmas GhassaniNo ratings yet
- Pellizzari 2008 - Gayralia SPP From Southern BrazilDocument8 pagesPellizzari 2008 - Gayralia SPP From Southern BrazilDaniellyNo ratings yet
- Breeding For Drought Tolerance in Tobacco PDFDocument10 pagesBreeding For Drought Tolerance in Tobacco PDFOgnen GaleskiNo ratings yet
- Physiological Characterization of Manzano Hot Pepper (Capsicum Pubescens R & P) LandracesDocument5 pagesPhysiological Characterization of Manzano Hot Pepper (Capsicum Pubescens R & P) LandracesLuis Saavedra Santa MariaNo ratings yet
- Agronomic and Phytochemical Characterization of CaiguaDocument6 pagesAgronomic and Phytochemical Characterization of Caiguaquiron333No ratings yet
- RECEPTIVIDADE ESTIGMÁTICA E VIABILIDADE POLÍNICA EM Theobroma SubincanumDocument7 pagesRECEPTIVIDADE ESTIGMÁTICA E VIABILIDADE POLÍNICA EM Theobroma SubincanumDaniela MoraisNo ratings yet
- Rcch,+01 Criollo Escobar+Moncayo Palacios+Lagos Burbano+2020+Phenology+and+Growth+of+Lulo+ (Solanum+Quitoense+Lam) +Plants+Grafted+Document10 pagesRcch,+01 Criollo Escobar+Moncayo Palacios+Lagos Burbano+2020+Phenology+and+Growth+of+Lulo+ (Solanum+Quitoense+Lam) +Plants+Grafted+Sofia Miku GutierrezNo ratings yet
- Variations Among Coffee (Coffea Arabica L.) Genotypes For Sensitivity To Drought Induced by Soil Drying at Early Stages of Growth in EthiopiaDocument10 pagesVariations Among Coffee (Coffea Arabica L.) Genotypes For Sensitivity To Drought Induced by Soil Drying at Early Stages of Growth in EthiopiaPremier PublishersNo ratings yet
- In Vitro Propagation of Blackberry (Rubus Fruticosus L.)Document12 pagesIn Vitro Propagation of Blackberry (Rubus Fruticosus L.)Keren Maciel100% (1)
- Mori 2016Document26 pagesMori 2016Luis OliveiraNo ratings yet
- Bharad 2009Document10 pagesBharad 2009Shri Hari PrasadNo ratings yet
- 2018 PrudenteDocument8 pages2018 PrudenteAntonioFlávioFerreiraNo ratings yet
- J E A P: Ournal of Nvironmental Nalysis and RogressDocument8 pagesJ E A P: Ournal of Nvironmental Nalysis and RogressGabriela MouraNo ratings yet
- Experimental re-WPS OfficeDocument10 pagesExperimental re-WPS OfficeChristian Dueña TabiosasNo ratings yet
- 4 FIOLOGI Document-DikonversiDocument34 pages4 FIOLOGI Document-DikonversiAsyahidah KhoirunnisaNo ratings yet
- Plant+Disease++Vol +94+no +8Document12 pagesPlant+Disease++Vol +94+no +8Virgilio UlloaNo ratings yet
- tmp22B5 TMPDocument9 pagestmp22B5 TMPFrontiersNo ratings yet
- Salinity Tolerance of Two Seedling Growth Stages of Annona Muricata L. in Port Harcourt, NigeriaDocument13 pagesSalinity Tolerance of Two Seedling Growth Stages of Annona Muricata L. in Port Harcourt, NigeriaCharityNo ratings yet
- Issn: Issn:: Acta Agronómica 0120-2812 2323-0118 Universidad Nacional de ColombiaDocument7 pagesIssn: Issn:: Acta Agronómica 0120-2812 2323-0118 Universidad Nacional de ColombiaYasunari arboleda hincapieNo ratings yet
- Evaluation of Potential Use of Native Anthurium Foliage: Echnical RticleDocument8 pagesEvaluation of Potential Use of Native Anthurium Foliage: Echnical RticleSilson CardosoNo ratings yet
- JPP Volume 3 Issue 1 Pages 109-116Document8 pagesJPP Volume 3 Issue 1 Pages 109-116bram samyNo ratings yet
- Effect of Host Plants On The Development, Survivorship, and Reproduction ofDocument8 pagesEffect of Host Plants On The Development, Survivorship, and Reproduction ofAndres ChuraNo ratings yet
- Microclimate, Development and Productivity of Robusta Coffee Shaded by Rubber Trees and at Full SunDocument10 pagesMicroclimate, Development and Productivity of Robusta Coffee Shaded by Rubber Trees and at Full SunPramedicaPerdanaPutraNo ratings yet
- Artigos em PeriódicoDocument6 pagesArtigos em PeriódicomarthancNo ratings yet
- Influence of Nitrogen and Phosphorus On The Growth and Yield of Cucumber (Cucumis Sativus L.)Document13 pagesInfluence of Nitrogen and Phosphorus On The Growth and Yield of Cucumber (Cucumis Sativus L.)Emmanuel Bassey EffaNo ratings yet
- 2454-Article Text-8029-1-10-20191022Document9 pages2454-Article Text-8029-1-10-20191022078 Medi Muhammad YusupNo ratings yet
- Metabolic and Physiologic Profile DuringDocument17 pagesMetabolic and Physiologic Profile DuringDaniel LDHSNo ratings yet
- Rosa Et. Al. 2021Document11 pagesRosa Et. Al. 2021Rosa Helena da SilvaNo ratings yet
- Araticum in Vitro SeedlingDocument4 pagesAraticum in Vitro Seedlingtcamel8No ratings yet
- Casestudy 15Document17 pagesCasestudy 15dawud kuroNo ratings yet
- FigueiredoEtMelo 2021b S New RecordsDocument7 pagesFigueiredoEtMelo 2021b S New RecordsgeorgesidneybarachoNo ratings yet
- Nitrogen and Potassium in Production, Nutrition and Water UseDocument10 pagesNitrogen and Potassium in Production, Nutrition and Water UseramdanNo ratings yet
- Garcia 2017Document8 pagesGarcia 2017Isidro Valencia Fertirriego MXNo ratings yet
- Absorcion de Nutrientes Del CrisantemoDocument10 pagesAbsorcion de Nutrientes Del CrisantemopectrinNo ratings yet
- Martins Da Costa Et Al 2017Document9 pagesMartins Da Costa Et Al 2017TamiSandobalNo ratings yet
- Chapter 9 PersonalityDocument66 pagesChapter 9 Personalitysp-hazaranaz.mohdaslamNo ratings yet
- BIOA01 F2021 SyllabusDocument10 pagesBIOA01 F2021 Syllabusthisis throwawayxDNo ratings yet
- Unnatural SelectionDocument2 pagesUnnatural SelectionkathleenNo ratings yet
- A Newsimplified ThermoregulatorybioheatmodelforevaluatingthermalDocument9 pagesA Newsimplified Thermoregulatorybioheatmodelforevaluatingthermalthi.rar3078No ratings yet
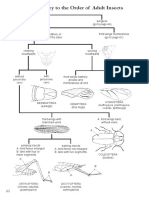
- Order Pictorial KeyDocument8 pagesOrder Pictorial KeyNova Tri Kusuma DewiNo ratings yet
- Genurile Aloinopsis, Nananthus Si Titanopsis PDFDocument34 pagesGenurile Aloinopsis, Nananthus Si Titanopsis PDFKty FloreaNo ratings yet
- Weiss JurmainDocument14 pagesWeiss JurmainLM AdrianneNo ratings yet
- Hypnosis Progressive Muscle Relaxation Induction ScriptDocument5 pagesHypnosis Progressive Muscle Relaxation Induction Scriptlezzo sudicioNo ratings yet
- Referat Engleza Tigrul AlbDocument2 pagesReferat Engleza Tigrul Albandrei0891No ratings yet
- GTSE - Online - Class V - EVSDocument13 pagesGTSE - Online - Class V - EVSranjanjhallbNo ratings yet
- Bio-Chemistry 2 MarksDocument49 pagesBio-Chemistry 2 Marksdevilcobra68No ratings yet
- Genetics Practice ProbsDocument4 pagesGenetics Practice Probsajaysharma19686191No ratings yet
- Bio CH4 F4 StudywithadminDocument16 pagesBio CH4 F4 StudywithadminRathi MalarNo ratings yet
- My Project New EditDocument38 pagesMy Project New Editmuhammad m butuNo ratings yet
- Soil-Plants-water RelationshipDocument10 pagesSoil-Plants-water RelationshipSunil DhankharNo ratings yet
- Gregory I. Bain, Eiji Itoi, Giovanni Di Giacomo, Hiroyuki Sugaya Eds. Normal and Pathological Anatomy of The ShoulderDocument410 pagesGregory I. Bain, Eiji Itoi, Giovanni Di Giacomo, Hiroyuki Sugaya Eds. Normal and Pathological Anatomy of The ShoulderNikolaos Anastasopoulos100% (4)
- Cell - 28 March 2013Document270 pagesCell - 28 March 2013Ian De La CruzNo ratings yet
- Daftar PustakaDocument5 pagesDaftar PustakaPramitha YustiaNo ratings yet
- GalactosemiaDocument41 pagesGalactosemiaArlene DaroNo ratings yet
- Child and AdolescentDocument2 pagesChild and AdolescentOrmon Angel AndesNo ratings yet
- Origin of Lymphoid Neoplasms: CLP: BLB: NBC: MC: GC: MZDocument26 pagesOrigin of Lymphoid Neoplasms: CLP: BLB: NBC: MC: GC: MZSri Naharindah NingNo ratings yet
- Evolutionary Artificial Intelligence Based Peptide Discoveries For EffectiveDocument13 pagesEvolutionary Artificial Intelligence Based Peptide Discoveries For EffectiveLucas PalmeiraNo ratings yet
- Anesthesiologists 26th AnnualMeeting New Orleans October 2001Document16 pagesAnesthesiologists 26th AnnualMeeting New Orleans October 2001Reginaldo CunhaNo ratings yet
- Research Work On Action of Homeopathic Medicines - DR Devendra Kumar MuntaDocument26 pagesResearch Work On Action of Homeopathic Medicines - DR Devendra Kumar MuntawdirectoryNo ratings yet
- Plant and Animal Cell OperationDocument8 pagesPlant and Animal Cell Operationapi-325015040No ratings yet
- Ascent of Sap by IDS PDFDocument27 pagesAscent of Sap by IDS PDFkingNo ratings yet
- Biol 111Document1 pageBiol 111Dammit DonNo ratings yet
- Guidance On Registration of Similar Biological Products in SingaporeDocument13 pagesGuidance On Registration of Similar Biological Products in SingaporeWilliam ChandraNo ratings yet
- Placenta Previa NCP 1Document6 pagesPlacenta Previa NCP 1Nicole ArandingNo ratings yet