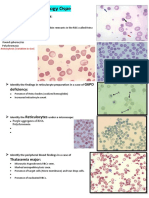
Download as pdf or txt
You might also like
- Practical Guide To Laboratory Test Interpretation and Differential Diagnosis. Haematology and BiochemistryDocument212 pagesPractical Guide To Laboratory Test Interpretation and Differential Diagnosis. Haematology and BiochemistryAgrovet Don Bosco100% (1)
- DXH Series Training Pocket Resources Spiralbook Rev A June2014Document124 pagesDXH Series Training Pocket Resources Spiralbook Rev A June2014dratultiwariNo ratings yet
- Erythrocyte Metabolism AND Membrane Structure and FunctionDocument29 pagesErythrocyte Metabolism AND Membrane Structure and FunctionJohn Marie IdavaNo ratings yet
- Membrane DeformabilityDocument2 pagesMembrane DeformabilityKenneth Jake BatiduanNo ratings yet
- RBC Membrane DeformabilityDocument16 pagesRBC Membrane DeformabilityMyedelle SeacorNo ratings yet
- Erythrocyte Metabolism and Membrane Structure and FunctionDocument38 pagesErythrocyte Metabolism and Membrane Structure and FunctionRej MaeNo ratings yet
- Plasma Membrane: The Lipid Bilayer Membrane ProteinsDocument79 pagesPlasma Membrane: The Lipid Bilayer Membrane ProteinsEarfanNo ratings yet
- Lesson 3 Red Cell Memebrane Developing-Metabolic PathwaysDocument19 pagesLesson 3 Red Cell Memebrane Developing-Metabolic Pathwaysتجربة أولىNo ratings yet
- Red Blood Cell BiochemistryDocument19 pagesRed Blood Cell BiochemistryPrincewill SeiyefaNo ratings yet
- Normal Erythropoiesis: A. Inner and Outer LayersDocument9 pagesNormal Erythropoiesis: A. Inner and Outer LayersTina MultazamiNo ratings yet
- HEMA LEC - ErythrocytesDocument5 pagesHEMA LEC - Erythrocytescherrycayari13No ratings yet
- Rbcs Metabolism: D.Hythem SaeedDocument19 pagesRbcs Metabolism: D.Hythem Saeedsamar yousif mohamedNo ratings yet
- Topic 3Document12 pagesTopic 3Ria AccadNo ratings yet
- Lect 1 Membrane Structure and FunctionDocument67 pagesLect 1 Membrane Structure and FunctionIzukanji kayoraNo ratings yet
- Module 3 - Lecture 3 - LectureDocument8 pagesModule 3 - Lecture 3 - LectureNi CaoNo ratings yet
- 01 Cell MembraneDocument22 pages01 Cell MembraneMian SahibNo ratings yet
- Chapter 4-Jnu Biomembranes-1 - 複本Document48 pagesChapter 4-Jnu Biomembranes-1 - 複本Wai Kwong ChiuNo ratings yet
- Lehninger PPT ch11 Wsuh 2024Document74 pagesLehninger PPT ch11 Wsuh 2024missoni0408No ratings yet
- RBC MembraneDocument41 pagesRBC MembranemwambungupNo ratings yet
- Chemical Composition of Blood Cells: MLS 302: Basic HaematologyDocument13 pagesChemical Composition of Blood Cells: MLS 302: Basic HaematologyBarakat IsmailNo ratings yet
- Fisiologi Cairan Tubuh & Darah: Dr. Nindya Aryanty, M.Med - EdDocument43 pagesFisiologi Cairan Tubuh & Darah: Dr. Nindya Aryanty, M.Med - EdIman TaufiqNo ratings yet
- EritrositDocument12 pagesEritrositNatasya HerinNo ratings yet
- 03lect Cell2 Summary2017Document4 pages03lect Cell2 Summary2017Dan ChoiNo ratings yet
- Cell Membrane (Structure and Function)Document150 pagesCell Membrane (Structure and Function)Dr. Tapan Kr. Dutta100% (1)
- Chapter 9 HemaDocument4 pagesChapter 9 HemaJoannah MarieNo ratings yet
- Study Exercise Biology 10-12 97.03Document7 pagesStudy Exercise Biology 10-12 97.03Fabiola Vania FeliciaNo ratings yet
- Athulya PlateletsDocument119 pagesAthulya Plateletsbetsy jonahsNo ratings yet
- Tissue BiochemistryDocument3 pagesTissue BiochemistryNeelam MehtabNo ratings yet
- Fishew1 Cell PhysiologyDocument27 pagesFishew1 Cell PhysiologyAdibMa'rufFauziNo ratings yet
- RBC MembraneDocument37 pagesRBC MembraneStanley ChikoveNo ratings yet
- K - 8 Blood Brain Barrier (Biokimia)Document31 pagesK - 8 Blood Brain Barrier (Biokimia)Firman YunusNo ratings yet
- 03 - The Structure of M. PlasmaDocument26 pages03 - The Structure of M. PlasmaZANUBA ULYA’ HAURA CELLIN 1No ratings yet
- Models of PlasmamembraneDocument19 pagesModels of PlasmamembraneGanesh V GaonkarNo ratings yet
- Lipid Traffic Floppy Drives and A SuperhighwayDocument12 pagesLipid Traffic Floppy Drives and A SuperhighwayJohan AlarcónNo ratings yet
- Chapter 5 Lipids Membrane LectureslidesDocument65 pagesChapter 5 Lipids Membrane Lectureslidesduy trầnNo ratings yet
- Erythropoiesis and AnaemiaDocument42 pagesErythropoiesis and AnaemiaKaylee NesbitNo ratings yet
- L03 Membrane StructureDocument37 pagesL03 Membrane StructureMa Christina Alessandra HingcoNo ratings yet
- The Cell MembraneDocument36 pagesThe Cell Membraneanulorance98No ratings yet
- Plasma MembraneDocument8 pagesPlasma Membranesakshisangwan02No ratings yet
- G Rojos BlancosDocument7 pagesG Rojos BlancosAndrea Valenzuela AedoNo ratings yet
- Plasma Membrane and Transport MechanismsDocument63 pagesPlasma Membrane and Transport MechanismsVaccedNo ratings yet
- 2 Cell Membrane 20210908Document83 pages2 Cell Membrane 20210908Sayan KonarNo ratings yet
- Lipid Movement BiochemDocument9 pagesLipid Movement BiochemCrowNo ratings yet
- Cell MembraneDocument44 pagesCell MembraneTere CantikkNo ratings yet
- Biochemistry & Genetics: II SHS 109: Resource Person: DR Tanveer Akbar Reference TextDocument60 pagesBiochemistry & Genetics: II SHS 109: Resource Person: DR Tanveer Akbar Reference TexttNo ratings yet
- Cell BiologyDocument41 pagesCell BiologyARJUN CHINMAYANo ratings yet
- 2.plasma MembraneDocument65 pages2.plasma MembraneAbdou BnelNo ratings yet
- Biologi CHP 6 Sem 1Document81 pagesBiologi CHP 6 Sem 1Aliaa AkbarNo ratings yet
- Biochemistry of Membrane LipidsDocument21 pagesBiochemistry of Membrane LipidsBhaskar GangulyNo ratings yet
- CellMembrane Structure and Functional PropertiesDocument5 pagesCellMembrane Structure and Functional PropertiesAnastasiafynnNo ratings yet
- Plasma Membrane in DetailsDocument24 pagesPlasma Membrane in DetailsSyeda Kaneez Mohammadi MohammadiNo ratings yet
- 8-Plasma Membranes 28-7-2020Document37 pages8-Plasma Membranes 28-7-2020Hiba N IkhmyesNo ratings yet
- ErytHroCyte ProduCtionDocument8 pagesErytHroCyte ProduCtionMichelle CaamicNo ratings yet
- BCH 419 NoteDocument20 pagesBCH 419 NoteJose BillNo ratings yet
- Intra Cellular Vesicular TrafficDocument48 pagesIntra Cellular Vesicular TrafficKanishkaNo ratings yet
- Membrane Structure and Function: Lecture OutlineDocument11 pagesMembrane Structure and Function: Lecture Outlinehaha_le12No ratings yet
- Bio 4.cellDocument59 pagesBio 4.cellAnish PandeyNo ratings yet
- BiomembranesDocument42 pagesBiomembranesChindhu SivasubramanianNo ratings yet
- Biology Molecular of Hematopoesis: Sry Suryani Widjaja Biochemistry Department September 2019Document47 pagesBiology Molecular of Hematopoesis: Sry Suryani Widjaja Biochemistry Department September 2019daffa fatih100% (1)
- Blood Cells Turnover PDFDocument4 pagesBlood Cells Turnover PDFNabillaMerdikaPutriKusumaNo ratings yet
- Red Blood Cells, Functions, Diseases A Simple Guide To The Condition, Diagnosis, Treatment, And Related ConditionsFrom EverandRed Blood Cells, Functions, Diseases A Simple Guide To The Condition, Diagnosis, Treatment, And Related ConditionsNo ratings yet
- Blood Anatomy and PhysiologyDocument4 pagesBlood Anatomy and PhysiologyRao Asad MubeenNo ratings yet
- (S.C. Rastogi) Essentials of Animal Physiology, 4t (BookSee - Org) 250Document1 page(S.C. Rastogi) Essentials of Animal Physiology, 4t (BookSee - Org) 250Indah Rizka AprilianiNo ratings yet
- ErythropoisesDocument30 pagesErythropoisesPriyanka RameshNo ratings yet
- Hematology PPT 1Document287 pagesHematology PPT 1TarishiMalikNo ratings yet
- Summary Assessment HISTO LABDocument8 pagesSummary Assessment HISTO LABARAH DE OCAMPONo ratings yet
- Biology MCQs For SSC ExamsDocument16 pagesBiology MCQs For SSC ExamsOrlinNo ratings yet
- Metals (PB, HG, MN, CD, As, Ni, CR)Document49 pagesMetals (PB, HG, MN, CD, As, Ni, CR)Йеша Маниш МираниNo ratings yet
- Case Presentation: Pre EclamsiaDocument66 pagesCase Presentation: Pre EclamsiaEugene Evan Endaya UyNo ratings yet
- Hemoglobin MetabolismDocument4 pagesHemoglobin Metabolismrona hilarioNo ratings yet
- Normal Erythrocyte1Document4 pagesNormal Erythrocyte1RonyArtoKapidaNo ratings yet
- HematologyDocument2 pagesHematologyMuhammad Hamza AlviNo ratings yet
- Basic HematologyDocument46 pagesBasic HematologySin RazyNo ratings yet
- Endobiosis or Blood Parasitism - The Teaching of Prof. G. EnderleinDocument47 pagesEndobiosis or Blood Parasitism - The Teaching of Prof. G. EnderleinrafaelNo ratings yet
- Erythrocyte Indices: JC Louise P. Bandala, RMTDocument18 pagesErythrocyte Indices: JC Louise P. Bandala, RMTLouise BandalaNo ratings yet
- Aplastic Hemolitic 2021 OlgaDocument43 pagesAplastic Hemolitic 2021 OlgalaibaNo ratings yet
- Hematopoiesis: Hematology 1Document10 pagesHematopoiesis: Hematology 1CdqdpNo ratings yet
- Question 1: Choose The Most Suitable Word or Phrase (A, B, C, or D) To Fill in Each Blank (20 Points)Document8 pagesQuestion 1: Choose The Most Suitable Word or Phrase (A, B, C, or D) To Fill in Each Blank (20 Points)Linh ChiNo ratings yet
- Hem Lab6 RBC Morph Incl F12Document7 pagesHem Lab6 RBC Morph Incl F12Elvan Dwi WidyadiNo ratings yet
- Platelet Diluting Fluid He892 PDFDocument1 pagePlatelet Diluting Fluid He892 PDFJohnree A. EvangelistaNo ratings yet
- URIT-3010 Operation ManualDocument90 pagesURIT-3010 Operation ManualzikossNo ratings yet
- Case Study Open FractureDocument56 pagesCase Study Open FractureEimereen Montebon Pileo100% (1)
- Hematology Ospe A1Document5 pagesHematology Ospe A1Zain Ul AbidinNo ratings yet
- Blood-Worksheet AP PDFDocument12 pagesBlood-Worksheet AP PDFShawnetz Copp100% (1)
- Format Laporan Praktikum Mikroteknik (Hewan) 2018 - Dion SatrioDocument9 pagesFormat Laporan Praktikum Mikroteknik (Hewan) 2018 - Dion SatrioDion Satrio PambudiNo ratings yet
- Physiology, Oxygen Transport: April 2019Document5 pagesPhysiology, Oxygen Transport: April 2019Soha SonaNo ratings yet
- Experiment #4: Demonstration of Common Immunohematologic ReactionsDocument5 pagesExperiment #4: Demonstration of Common Immunohematologic ReactionsKriziaNo ratings yet
- Pathology Question BankDocument10 pagesPathology Question BankSana UsmaniNo ratings yet
- BloodDocument18 pagesBloodadnankhaNo ratings yet