
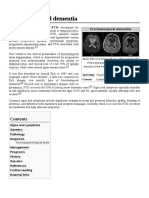
RohrerCON2011
RohrerCON2011
You might also like
- Uworld Charts PDFDocument162 pagesUworld Charts PDFMaryam Fadah100% (3)
- SBM WINS HYGIENE Narrative Report 2019 FinalDocument12 pagesSBM WINS HYGIENE Narrative Report 2019 FinalPauline Intia83% (30)
- 17A. Immune ToleranceDocument8 pages17A. Immune ToleranceEssington BeloNo ratings yet
- 19 Literature Review - Primary MyelofibrosisDocument10 pages19 Literature Review - Primary MyelofibrosisMonica CalisayaNo ratings yet
- Frontotemporal Dementia NeuropathologyDocument14 pagesFrontotemporal Dementia Neuropathologyppico1963No ratings yet
- Sun Et Al 2019Document5 pagesSun Et Al 2019MiaNo ratings yet
- 1 - Investigação Genética Das Demências Na Prática ClínicaDocument6 pages1 - Investigação Genética Das Demências Na Prática Clínicaveidamororo17No ratings yet
- 67 - Biparietal Variant of Alzheimer's Disease A Rare Presentation of A Common DiseaseDocument17 pages67 - Biparietal Variant of Alzheimer's Disease A Rare Presentation of A Common DiseaseFaras ArinalNo ratings yet
- GeneticsDocument10 pagesGeneticsapi-3835615No ratings yet
- znl475Document9 pagesznl475m del mar cantón neffNo ratings yet
- Frontotemporal Dementia An Updated Overview.14Document5 pagesFrontotemporal Dementia An Updated Overview.1425paramveerNo ratings yet
- 61-1-13sd RettDocument11 pages61-1-13sd RettCarolina GuzmánNo ratings yet
- Journal - Collection of Journal NEUROCHEMISTRY 2006Document10 pagesJournal - Collection of Journal NEUROCHEMISTRY 2006leozdmNo ratings yet
- s41380 021 01384 8Document10 pagess41380 021 01384 8mariamojegNo ratings yet
- 2023 What A Neurologist Must Know of TTR AmyloidosisDocument11 pages2023 What A Neurologist Must Know of TTR AmyloidosisAlejandra Gonzalez DuarteNo ratings yet
- Neuroblastoma: Biological Insights Into A Clinical Enigma: Garrett M. BrodeurDocument14 pagesNeuroblastoma: Biological Insights Into A Clinical Enigma: Garrett M. BrodeurMouris DwiputraNo ratings yet
- Fronto Temporal Dementia - FullDocument12 pagesFronto Temporal Dementia - FullnowheretidelandNo ratings yet
- Frontotemporal Dementia: Diagnosis, Deficits and Management: ReviewDocument16 pagesFrontotemporal Dementia: Diagnosis, Deficits and Management: ReviewGislaine GuerraNo ratings yet
- The Landscape of Epilepsy-Related GATOR1 Variants Baldassari 2019Document11 pagesThe Landscape of Epilepsy-Related GATOR1 Variants Baldassari 2019cicasmileNo ratings yet
- Progranulin in Neurodegenerative PDFDocument11 pagesProgranulin in Neurodegenerative PDFlnb71freeNo ratings yet
- Clinicopathological Features of The Tauopathies - MUrray - MolMechNeurodeg - 2005Document5 pagesClinicopathological Features of The Tauopathies - MUrray - MolMechNeurodeg - 2005MARIA MONTSERRAT SOMOZA MONCADANo ratings yet
- 2013 - Martin and Wong - Aberrant Regulation of DNA Methylation in Amyotrophic Lateral Sclerosis A New Target of Disease MechanismsDocument12 pages2013 - Martin and Wong - Aberrant Regulation of DNA Methylation in Amyotrophic Lateral Sclerosis A New Target of Disease Mechanismsmarej312No ratings yet
- Mitochondrial Disorders OverviewDocument21 pagesMitochondrial Disorders OverviewIt's MeNo ratings yet
- Mitochondrial Encephalomyopathies (MEM)Document66 pagesMitochondrial Encephalomyopathies (MEM)Fatma KaledNo ratings yet
- Linfoma Folicular com Lesão em BocaDocument5 pagesLinfoma Folicular com Lesão em Bocagsaraujo1990No ratings yet
- Vocal Cord Paresis and Diaphragmatic DysDocument11 pagesVocal Cord Paresis and Diaphragmatic DysMILON KUMAR HORENo ratings yet
- Molecular and Cellular Neuroscience: Petra Steinacker, Peggy Barschke, Markus Otto TDocument17 pagesMolecular and Cellular Neuroscience: Petra Steinacker, Peggy Barschke, Markus Otto Tfazila zahoorNo ratings yet
- Frontotemporal DementiaDocument11 pagesFrontotemporal DementiaJussel Vazquez MarquezNo ratings yet
- Npjamd 20153Document11 pagesNpjamd 20153Marija PetrovicNo ratings yet
- Fragile X Syndrome: Causes, Diagnosis, Mechanisms, and TherapeuticsDocument9 pagesFragile X Syndrome: Causes, Diagnosis, Mechanisms, and TherapeuticshanzelNo ratings yet
- Neurofibromatosis Type 1: Ontinuing Medical EducationDocument14 pagesNeurofibromatosis Type 1: Ontinuing Medical EducationMatthew DavisNo ratings yet
- Novel Frizzled-4 Gene Mutations in Chinese Patients With Familial Exudative VitreoretinopathyDocument9 pagesNovel Frizzled-4 Gene Mutations in Chinese Patients With Familial Exudative VitreoretinopathyMiaNo ratings yet
- Fneur 12 648740Document7 pagesFneur 12 648740june.talon7No ratings yet
- Unstable Expansion of Triplet Repeats As A New Disease Mechanism For Neurodegenerative DiseasesDocument12 pagesUnstable Expansion of Triplet Repeats As A New Disease Mechanism For Neurodegenerative Diseasestkovats.lopes3No ratings yet
- Mitochondrial Disorders Overview - GeneReviews® - NCBI Bookshelf HTTPSWWW - Ncbi.nlm - nih.govbooksNBK1224Document24 pagesMitochondrial Disorders Overview - GeneReviews® - NCBI Bookshelf HTTPSWWW - Ncbi.nlm - nih.govbooksNBK1224denisaNo ratings yet
- Man 2002Document9 pagesMan 2002GatoNo ratings yet
- Novel Insight Into Intellectual Disability A RevieDocument9 pagesNovel Insight Into Intellectual Disability A RevieJuan Jose Eraso OsorioNo ratings yet
- TRIVLI Et Al. 2020. Primary Open Angle Glaucoma Genetics, The Common Variants and Their Clinical Associations (Review)Document8 pagesTRIVLI Et Al. 2020. Primary Open Angle Glaucoma Genetics, The Common Variants and Their Clinical Associations (Review)Thiago RodriguesNo ratings yet
- NEJMoa 074306Document10 pagesNEJMoa 074306monamustafaNo ratings yet
- Zs Urka 2013Document4 pagesZs Urka 2013trukopNo ratings yet
- Dementia Is A Syndrome Characterized by An Appreciable Deterioration in Cognition Resulting in Behavioral Problems andDocument2 pagesDementia Is A Syndrome Characterized by An Appreciable Deterioration in Cognition Resulting in Behavioral Problems andAllandale NacinoNo ratings yet
- Sobrelapamiento ELA y DFT 2020Document10 pagesSobrelapamiento ELA y DFT 2020siralkNo ratings yet
- Cooper-14 C9orf72 Espectro ClínicoDocument13 pagesCooper-14 C9orf72 Espectro ClínicoMARIA MONTSERRAT SOMOZA MONCADANo ratings yet
- edbook_am.2012.32.85Document8 pagesedbook_am.2012.32.85Munazza RashidNo ratings yet
- Genetic Association Study of Putative Functional Single Nucleotide Polymorphisms of Genes in Folate Metabolism and Spina BifidaDocument11 pagesGenetic Association Study of Putative Functional Single Nucleotide Polymorphisms of Genes in Folate Metabolism and Spina BifidaMauro Porcel de PeraltaNo ratings yet
- Duplication at Xq13.3-q21.1 With Syndromic Intellectual Disability, A Probable Role For The ATRX GeneDocument6 pagesDuplication at Xq13.3-q21.1 With Syndromic Intellectual Disability, A Probable Role For The ATRX GeneEyyup UctepeNo ratings yet
- DistoniaDocument5 pagesDistoniaMarlon RojasNo ratings yet
- VIP Cives Oncotarget 2016Document16 pagesVIP Cives Oncotarget 2016Jemma ArakelyanNo ratings yet
- ddu440 jph1 ER-mito-CaDocument17 pagesddu440 jph1 ER-mito-CaJaimeD.LópezAlcaláNo ratings yet
- Abstract BookDocument90 pagesAbstract BookPopescu RoxanaNo ratings yet
- 2011 Dickson FTLD TauDocument12 pages2011 Dickson FTLD TauvkarmoNo ratings yet
- Clinical and Genetic Basis of Familial Amyotrophic Lateral Sclerosis (Revisión)Document12 pagesClinical and Genetic Basis of Familial Amyotrophic Lateral Sclerosis (Revisión)Francisco Ahumada MéndezNo ratings yet
- Wu Et Al 2016Document11 pagesWu Et Al 2016MiaNo ratings yet
- Polycystic Kidney - Eight CasesDocument11 pagesPolycystic Kidney - Eight CasesKAINAT ANJUMNo ratings yet
- Patterns of Genetic Transmission & Hereditary Hearing Loss: DR - DivyaDocument60 pagesPatterns of Genetic Transmission & Hereditary Hearing Loss: DR - DivyavipinNo ratings yet
- Seminar: Anthony H V SchapiraDocument10 pagesSeminar: Anthony H V Schapiramompou88No ratings yet
- Phenotype-genotype Relations in FTDDocument19 pagesPhenotype-genotype Relations in FTDKeerthana SomashekarNo ratings yet
- E74 FullDocument5 pagesE74 FulldhiaNo ratings yet
- Fragile X and GeneticsDocument8 pagesFragile X and GeneticsS ANo ratings yet
- Nihms 650938Document19 pagesNihms 650938chetanNo ratings yet
- 2012 JNeurol Neurosurg Psy Rossor CMTreviewDocument11 pages2012 JNeurol Neurosurg Psy Rossor CMTreviewDr Waseem RajaNo ratings yet
- Epidemiology, Natural History, Pathology and Management of Medulloblastoma in ChildrenDocument48 pagesEpidemiology, Natural History, Pathology and Management of Medulloblastoma in ChildrenSam OlukaNo ratings yet
- Auto-Inflammatory Syndromes: Pathophysiology, Diagnosis, and ManagementFrom EverandAuto-Inflammatory Syndromes: Pathophysiology, Diagnosis, and ManagementPetros EfthimiouNo ratings yet
- Vocabulary List 1: Cyril's Most Impressive Quality Was His Innate Goodness. He Had An Innate MusicalityDocument28 pagesVocabulary List 1: Cyril's Most Impressive Quality Was His Innate Goodness. He Had An Innate MusicalityYuk Ngo FongNo ratings yet
- Cucumber As An Organic Ingredient For Laundry Detergent FINALDocument49 pagesCucumber As An Organic Ingredient For Laundry Detergent FINALIwa100% (1)
- Mesentery, Omentum, Peritoneum: Inflammatory, Infectious Diseases and Pseudo LesionsDocument11 pagesMesentery, Omentum, Peritoneum: Inflammatory, Infectious Diseases and Pseudo LesionsRendra SyaniNo ratings yet
- Docsbookstoreacsapac2022b1 Sample PDFDocument30 pagesDocsbookstoreacsapac2022b1 Sample PDFFaheem Mahmood ButtNo ratings yet
- Role of Ophthalmic Nurses in Prevention of Ophthalmic DiseasesDocument4 pagesRole of Ophthalmic Nurses in Prevention of Ophthalmic DiseaseskaskwawNo ratings yet
- Personal Support Worker NBDocument5 pagesPersonal Support Worker NBOgunsakin OluwaseyiNo ratings yet
- HypotoniaDocument45 pagesHypotoniaاثير اااNo ratings yet
- Immune Cell Tracking in Vivo (IVIS) Using BODIPY DyeDocument5 pagesImmune Cell Tracking in Vivo (IVIS) Using BODIPY DyeJamieNo ratings yet
- Katkam 2016Document7 pagesKatkam 2016Thishya KalluriNo ratings yet
- Neurology Coe MDocument39 pagesNeurology Coe MSalviandy DhanizarNo ratings yet
- BUDGET OF WORK (AutoRecovered)Document10 pagesBUDGET OF WORK (AutoRecovered)Romel GarciaNo ratings yet
- Exam Program Oct 2022 Ctle (Chem) - FinalDocument4 pagesExam Program Oct 2022 Ctle (Chem) - Finalraymond tuizaNo ratings yet
- Psyc Chapter 5Document18 pagesPsyc Chapter 5lordessofdawnNo ratings yet
- World Health DayDocument2 pagesWorld Health DaySahilNo ratings yet
- NRPB W67Document107 pagesNRPB W67Cecilia BarabinoNo ratings yet
- EIA M A: OF Aleic NhydrideDocument7 pagesEIA M A: OF Aleic NhydrideNoman AslamNo ratings yet
- Textbook Mayo Clinic Internal Medicine Board Review 11 Edition Edition Christopher M Wittich Ebook All Chapter PDFDocument53 pagesTextbook Mayo Clinic Internal Medicine Board Review 11 Edition Edition Christopher M Wittich Ebook All Chapter PDFsharon.janssen791100% (13)
- Chapter 13 Rodaks HematologyDocument10 pagesChapter 13 Rodaks HematologyRALPH JAN T. RIONo ratings yet
- Principles of Patient'S Safety: Basic Health Skills (GRU1251)Document39 pagesPrinciples of Patient'S Safety: Basic Health Skills (GRU1251)attooNo ratings yet
- ASchool-BasedStudyoftheInfluenceofStudentsRelationshipAPJCP Volume19 IssueS1 Pages7-121Document7 pagesASchool-BasedStudyoftheInfluenceofStudentsRelationshipAPJCP Volume19 IssueS1 Pages7-121king12No ratings yet
- ABC Xanthoxylum FraxineumDocument43 pagesABC Xanthoxylum Fraxineumrafeek88pmNo ratings yet
- Understanding The Mechanism of Chikungunya Virus Vector Competence in Three Species of MosquitoesDocument13 pagesUnderstanding The Mechanism of Chikungunya Virus Vector Competence in Three Species of MosquitoesRayssa OliveiraNo ratings yet
- English Remidial/Enrichment: Nama: Aisi Kurniasari Kelas: 12 Tata Busana 3 No. Absen: 01Document8 pagesEnglish Remidial/Enrichment: Nama: Aisi Kurniasari Kelas: 12 Tata Busana 3 No. Absen: 01Tlebeng AstreaNo ratings yet
- Pelvic Inflammatory Disease - EditedDocument7 pagesPelvic Inflammatory Disease - EditedDanNo ratings yet
- Drug Facts: ALLER-CHLOR-chlorpheniramine Maleate S Yrup Rugby LaboratoriesDocument4 pagesDrug Facts: ALLER-CHLOR-chlorpheniramine Maleate S Yrup Rugby LaboratoriesAsif KhanNo ratings yet
- Medical Form 2024Document2 pagesMedical Form 2024shellyann brightly-gordonNo ratings yet
- 118 Legal and Critical ReviewerDocument5 pages118 Legal and Critical ReviewerJeminah Fae CalmaNo ratings yet























































































Download as pdf or txt
You might also like
- Uworld Charts PDFDocument162 pagesUworld Charts PDFMaryam Fadah100% (3)
- SBM WINS HYGIENE Narrative Report 2019 FinalDocument12 pagesSBM WINS HYGIENE Narrative Report 2019 FinalPauline Intia83% (30)
- 17A. Immune ToleranceDocument8 pages17A. Immune ToleranceEssington BeloNo ratings yet
- 19 Literature Review - Primary MyelofibrosisDocument10 pages19 Literature Review - Primary MyelofibrosisMonica CalisayaNo ratings yet
- Frontotemporal Dementia NeuropathologyDocument14 pagesFrontotemporal Dementia Neuropathologyppico1963No ratings yet
- Sun Et Al 2019Document5 pagesSun Et Al 2019MiaNo ratings yet
- 1 - Investigação Genética Das Demências Na Prática ClínicaDocument6 pages1 - Investigação Genética Das Demências Na Prática Clínicaveidamororo17No ratings yet
- 67 - Biparietal Variant of Alzheimer's Disease A Rare Presentation of A Common DiseaseDocument17 pages67 - Biparietal Variant of Alzheimer's Disease A Rare Presentation of A Common DiseaseFaras ArinalNo ratings yet
- GeneticsDocument10 pagesGeneticsapi-3835615No ratings yet
- znl475Document9 pagesznl475m del mar cantón neffNo ratings yet
- Frontotemporal Dementia An Updated Overview.14Document5 pagesFrontotemporal Dementia An Updated Overview.1425paramveerNo ratings yet
- 61-1-13sd RettDocument11 pages61-1-13sd RettCarolina GuzmánNo ratings yet
- Journal - Collection of Journal NEUROCHEMISTRY 2006Document10 pagesJournal - Collection of Journal NEUROCHEMISTRY 2006leozdmNo ratings yet
- s41380 021 01384 8Document10 pagess41380 021 01384 8mariamojegNo ratings yet
- 2023 What A Neurologist Must Know of TTR AmyloidosisDocument11 pages2023 What A Neurologist Must Know of TTR AmyloidosisAlejandra Gonzalez DuarteNo ratings yet
- Neuroblastoma: Biological Insights Into A Clinical Enigma: Garrett M. BrodeurDocument14 pagesNeuroblastoma: Biological Insights Into A Clinical Enigma: Garrett M. BrodeurMouris DwiputraNo ratings yet
- Fronto Temporal Dementia - FullDocument12 pagesFronto Temporal Dementia - FullnowheretidelandNo ratings yet
- Frontotemporal Dementia: Diagnosis, Deficits and Management: ReviewDocument16 pagesFrontotemporal Dementia: Diagnosis, Deficits and Management: ReviewGislaine GuerraNo ratings yet
- The Landscape of Epilepsy-Related GATOR1 Variants Baldassari 2019Document11 pagesThe Landscape of Epilepsy-Related GATOR1 Variants Baldassari 2019cicasmileNo ratings yet
- Progranulin in Neurodegenerative PDFDocument11 pagesProgranulin in Neurodegenerative PDFlnb71freeNo ratings yet
- Clinicopathological Features of The Tauopathies - MUrray - MolMechNeurodeg - 2005Document5 pagesClinicopathological Features of The Tauopathies - MUrray - MolMechNeurodeg - 2005MARIA MONTSERRAT SOMOZA MONCADANo ratings yet
- 2013 - Martin and Wong - Aberrant Regulation of DNA Methylation in Amyotrophic Lateral Sclerosis A New Target of Disease MechanismsDocument12 pages2013 - Martin and Wong - Aberrant Regulation of DNA Methylation in Amyotrophic Lateral Sclerosis A New Target of Disease Mechanismsmarej312No ratings yet
- Mitochondrial Disorders OverviewDocument21 pagesMitochondrial Disorders OverviewIt's MeNo ratings yet
- Mitochondrial Encephalomyopathies (MEM)Document66 pagesMitochondrial Encephalomyopathies (MEM)Fatma KaledNo ratings yet
- Linfoma Folicular com Lesão em BocaDocument5 pagesLinfoma Folicular com Lesão em Bocagsaraujo1990No ratings yet
- Vocal Cord Paresis and Diaphragmatic DysDocument11 pagesVocal Cord Paresis and Diaphragmatic DysMILON KUMAR HORENo ratings yet
- Molecular and Cellular Neuroscience: Petra Steinacker, Peggy Barschke, Markus Otto TDocument17 pagesMolecular and Cellular Neuroscience: Petra Steinacker, Peggy Barschke, Markus Otto Tfazila zahoorNo ratings yet
- Frontotemporal DementiaDocument11 pagesFrontotemporal DementiaJussel Vazquez MarquezNo ratings yet
- Npjamd 20153Document11 pagesNpjamd 20153Marija PetrovicNo ratings yet
- Fragile X Syndrome: Causes, Diagnosis, Mechanisms, and TherapeuticsDocument9 pagesFragile X Syndrome: Causes, Diagnosis, Mechanisms, and TherapeuticshanzelNo ratings yet
- Neurofibromatosis Type 1: Ontinuing Medical EducationDocument14 pagesNeurofibromatosis Type 1: Ontinuing Medical EducationMatthew DavisNo ratings yet
- Novel Frizzled-4 Gene Mutations in Chinese Patients With Familial Exudative VitreoretinopathyDocument9 pagesNovel Frizzled-4 Gene Mutations in Chinese Patients With Familial Exudative VitreoretinopathyMiaNo ratings yet
- Fneur 12 648740Document7 pagesFneur 12 648740june.talon7No ratings yet
- Unstable Expansion of Triplet Repeats As A New Disease Mechanism For Neurodegenerative DiseasesDocument12 pagesUnstable Expansion of Triplet Repeats As A New Disease Mechanism For Neurodegenerative Diseasestkovats.lopes3No ratings yet
- Mitochondrial Disorders Overview - GeneReviews® - NCBI Bookshelf HTTPSWWW - Ncbi.nlm - nih.govbooksNBK1224Document24 pagesMitochondrial Disorders Overview - GeneReviews® - NCBI Bookshelf HTTPSWWW - Ncbi.nlm - nih.govbooksNBK1224denisaNo ratings yet
- Man 2002Document9 pagesMan 2002GatoNo ratings yet
- Novel Insight Into Intellectual Disability A RevieDocument9 pagesNovel Insight Into Intellectual Disability A RevieJuan Jose Eraso OsorioNo ratings yet
- TRIVLI Et Al. 2020. Primary Open Angle Glaucoma Genetics, The Common Variants and Their Clinical Associations (Review)Document8 pagesTRIVLI Et Al. 2020. Primary Open Angle Glaucoma Genetics, The Common Variants and Their Clinical Associations (Review)Thiago RodriguesNo ratings yet
- NEJMoa 074306Document10 pagesNEJMoa 074306monamustafaNo ratings yet
- Zs Urka 2013Document4 pagesZs Urka 2013trukopNo ratings yet
- Dementia Is A Syndrome Characterized by An Appreciable Deterioration in Cognition Resulting in Behavioral Problems andDocument2 pagesDementia Is A Syndrome Characterized by An Appreciable Deterioration in Cognition Resulting in Behavioral Problems andAllandale NacinoNo ratings yet
- Sobrelapamiento ELA y DFT 2020Document10 pagesSobrelapamiento ELA y DFT 2020siralkNo ratings yet
- Cooper-14 C9orf72 Espectro ClínicoDocument13 pagesCooper-14 C9orf72 Espectro ClínicoMARIA MONTSERRAT SOMOZA MONCADANo ratings yet
- edbook_am.2012.32.85Document8 pagesedbook_am.2012.32.85Munazza RashidNo ratings yet
- Genetic Association Study of Putative Functional Single Nucleotide Polymorphisms of Genes in Folate Metabolism and Spina BifidaDocument11 pagesGenetic Association Study of Putative Functional Single Nucleotide Polymorphisms of Genes in Folate Metabolism and Spina BifidaMauro Porcel de PeraltaNo ratings yet
- Duplication at Xq13.3-q21.1 With Syndromic Intellectual Disability, A Probable Role For The ATRX GeneDocument6 pagesDuplication at Xq13.3-q21.1 With Syndromic Intellectual Disability, A Probable Role For The ATRX GeneEyyup UctepeNo ratings yet
- DistoniaDocument5 pagesDistoniaMarlon RojasNo ratings yet
- VIP Cives Oncotarget 2016Document16 pagesVIP Cives Oncotarget 2016Jemma ArakelyanNo ratings yet
- ddu440 jph1 ER-mito-CaDocument17 pagesddu440 jph1 ER-mito-CaJaimeD.LópezAlcaláNo ratings yet
- Abstract BookDocument90 pagesAbstract BookPopescu RoxanaNo ratings yet
- 2011 Dickson FTLD TauDocument12 pages2011 Dickson FTLD TauvkarmoNo ratings yet
- Clinical and Genetic Basis of Familial Amyotrophic Lateral Sclerosis (Revisión)Document12 pagesClinical and Genetic Basis of Familial Amyotrophic Lateral Sclerosis (Revisión)Francisco Ahumada MéndezNo ratings yet
- Wu Et Al 2016Document11 pagesWu Et Al 2016MiaNo ratings yet
- Polycystic Kidney - Eight CasesDocument11 pagesPolycystic Kidney - Eight CasesKAINAT ANJUMNo ratings yet
- Patterns of Genetic Transmission & Hereditary Hearing Loss: DR - DivyaDocument60 pagesPatterns of Genetic Transmission & Hereditary Hearing Loss: DR - DivyavipinNo ratings yet
- Seminar: Anthony H V SchapiraDocument10 pagesSeminar: Anthony H V Schapiramompou88No ratings yet
- Phenotype-genotype Relations in FTDDocument19 pagesPhenotype-genotype Relations in FTDKeerthana SomashekarNo ratings yet
- E74 FullDocument5 pagesE74 FulldhiaNo ratings yet
- Fragile X and GeneticsDocument8 pagesFragile X and GeneticsS ANo ratings yet
- Nihms 650938Document19 pagesNihms 650938chetanNo ratings yet
- 2012 JNeurol Neurosurg Psy Rossor CMTreviewDocument11 pages2012 JNeurol Neurosurg Psy Rossor CMTreviewDr Waseem RajaNo ratings yet
- Epidemiology, Natural History, Pathology and Management of Medulloblastoma in ChildrenDocument48 pagesEpidemiology, Natural History, Pathology and Management of Medulloblastoma in ChildrenSam OlukaNo ratings yet
- Auto-Inflammatory Syndromes: Pathophysiology, Diagnosis, and ManagementFrom EverandAuto-Inflammatory Syndromes: Pathophysiology, Diagnosis, and ManagementPetros EfthimiouNo ratings yet
- Vocabulary List 1: Cyril's Most Impressive Quality Was His Innate Goodness. He Had An Innate MusicalityDocument28 pagesVocabulary List 1: Cyril's Most Impressive Quality Was His Innate Goodness. He Had An Innate MusicalityYuk Ngo FongNo ratings yet
- Cucumber As An Organic Ingredient For Laundry Detergent FINALDocument49 pagesCucumber As An Organic Ingredient For Laundry Detergent FINALIwa100% (1)
- Mesentery, Omentum, Peritoneum: Inflammatory, Infectious Diseases and Pseudo LesionsDocument11 pagesMesentery, Omentum, Peritoneum: Inflammatory, Infectious Diseases and Pseudo LesionsRendra SyaniNo ratings yet
- Docsbookstoreacsapac2022b1 Sample PDFDocument30 pagesDocsbookstoreacsapac2022b1 Sample PDFFaheem Mahmood ButtNo ratings yet
- Role of Ophthalmic Nurses in Prevention of Ophthalmic DiseasesDocument4 pagesRole of Ophthalmic Nurses in Prevention of Ophthalmic DiseaseskaskwawNo ratings yet
- Personal Support Worker NBDocument5 pagesPersonal Support Worker NBOgunsakin OluwaseyiNo ratings yet
- HypotoniaDocument45 pagesHypotoniaاثير اااNo ratings yet
- Immune Cell Tracking in Vivo (IVIS) Using BODIPY DyeDocument5 pagesImmune Cell Tracking in Vivo (IVIS) Using BODIPY DyeJamieNo ratings yet
- Katkam 2016Document7 pagesKatkam 2016Thishya KalluriNo ratings yet
- Neurology Coe MDocument39 pagesNeurology Coe MSalviandy DhanizarNo ratings yet
- BUDGET OF WORK (AutoRecovered)Document10 pagesBUDGET OF WORK (AutoRecovered)Romel GarciaNo ratings yet
- Exam Program Oct 2022 Ctle (Chem) - FinalDocument4 pagesExam Program Oct 2022 Ctle (Chem) - Finalraymond tuizaNo ratings yet
- Psyc Chapter 5Document18 pagesPsyc Chapter 5lordessofdawnNo ratings yet
- World Health DayDocument2 pagesWorld Health DaySahilNo ratings yet
- NRPB W67Document107 pagesNRPB W67Cecilia BarabinoNo ratings yet
- EIA M A: OF Aleic NhydrideDocument7 pagesEIA M A: OF Aleic NhydrideNoman AslamNo ratings yet
- Textbook Mayo Clinic Internal Medicine Board Review 11 Edition Edition Christopher M Wittich Ebook All Chapter PDFDocument53 pagesTextbook Mayo Clinic Internal Medicine Board Review 11 Edition Edition Christopher M Wittich Ebook All Chapter PDFsharon.janssen791100% (13)
- Chapter 13 Rodaks HematologyDocument10 pagesChapter 13 Rodaks HematologyRALPH JAN T. RIONo ratings yet
- Principles of Patient'S Safety: Basic Health Skills (GRU1251)Document39 pagesPrinciples of Patient'S Safety: Basic Health Skills (GRU1251)attooNo ratings yet
- ASchool-BasedStudyoftheInfluenceofStudentsRelationshipAPJCP Volume19 IssueS1 Pages7-121Document7 pagesASchool-BasedStudyoftheInfluenceofStudentsRelationshipAPJCP Volume19 IssueS1 Pages7-121king12No ratings yet
- ABC Xanthoxylum FraxineumDocument43 pagesABC Xanthoxylum Fraxineumrafeek88pmNo ratings yet
- Understanding The Mechanism of Chikungunya Virus Vector Competence in Three Species of MosquitoesDocument13 pagesUnderstanding The Mechanism of Chikungunya Virus Vector Competence in Three Species of MosquitoesRayssa OliveiraNo ratings yet
- English Remidial/Enrichment: Nama: Aisi Kurniasari Kelas: 12 Tata Busana 3 No. Absen: 01Document8 pagesEnglish Remidial/Enrichment: Nama: Aisi Kurniasari Kelas: 12 Tata Busana 3 No. Absen: 01Tlebeng AstreaNo ratings yet
- Pelvic Inflammatory Disease - EditedDocument7 pagesPelvic Inflammatory Disease - EditedDanNo ratings yet
- Drug Facts: ALLER-CHLOR-chlorpheniramine Maleate S Yrup Rugby LaboratoriesDocument4 pagesDrug Facts: ALLER-CHLOR-chlorpheniramine Maleate S Yrup Rugby LaboratoriesAsif KhanNo ratings yet
- Medical Form 2024Document2 pagesMedical Form 2024shellyann brightly-gordonNo ratings yet
- 118 Legal and Critical ReviewerDocument5 pages118 Legal and Critical ReviewerJeminah Fae CalmaNo ratings yet