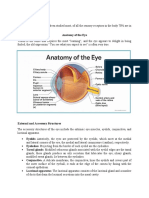
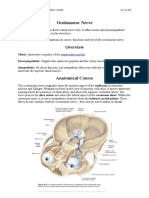
Download as rtf, pdf, or txt
You might also like
- Test Bank For Medical Assisting Administrative and Clinical Competencies 8th Edition by BlesiDocument10 pagesTest Bank For Medical Assisting Administrative and Clinical Competencies 8th Edition by Blesisophiacarrdabemqozri100% (26)
- 234 Coordinated Eye MovementsDocument1 page234 Coordinated Eye MovementsVasilii LentitchiiNo ratings yet
- 236 Coordinated Eye MovementsDocument1 page236 Coordinated Eye MovementsVasilii LentitchiiNo ratings yet
- Assignment 1 Year 3Document52 pagesAssignment 1 Year 3Tryphosa MalamboNo ratings yet
- Physiology of Vision-Sem4-Special SenseDocument20 pagesPhysiology of Vision-Sem4-Special SenseYUSMAWATI YUSRANNo ratings yet
- Visual and AuditoryDocument7 pagesVisual and AuditoryChristi EspinosaNo ratings yet
- Eye MovementsDocument18 pagesEye MovementsVaz CaNo ratings yet
- Chapter 8 Ocular Motor ControlDocument1 pageChapter 8 Ocular Motor ControlDeena Al NuaimiNo ratings yet
- Visual Neuroscience Revision QuestionsDocument6 pagesVisual Neuroscience Revision QuestionsAlex MNo ratings yet
- BLOK 1.3 Jaras PenglihatanDocument43 pagesBLOK 1.3 Jaras PenglihatanlenypsNo ratings yet
- Cooperation of The EyeDocument5 pagesCooperation of The EyeAmi MisiciNo ratings yet
- Perception of Moevemmt PDFDocument6 pagesPerception of Moevemmt PDFAbrar AhmadNo ratings yet
- Cranial NervesDocument61 pagesCranial NervesGuo YageNo ratings yet
- Medical Neuroscience Tutorial Notes: Vestibular System-Central ProcessingDocument3 pagesMedical Neuroscience Tutorial Notes: Vestibular System-Central ProcessingHenrique De Figueiredo MariscoNo ratings yet
- Neuroanatomy, Pupillary Light Reflexes and PathwayDocument6 pagesNeuroanatomy, Pupillary Light Reflexes and PathwayMatthew PrestonNo ratings yet
- Physiology of BalanceDocument45 pagesPhysiology of BalanceThadei MrumaNo ratings yet
- The Primary Visual CortexDocument30 pagesThe Primary Visual CortexΕιρήνηΔασκιωτάκηNo ratings yet
- Anatomy of The EyeDocument4 pagesAnatomy of The EyeSabrinaAyuPutriNo ratings yet
- Sheena The EyeDocument9 pagesSheena The EyeSheena Yen de Pano-PagdalianNo ratings yet
- Human SensesDocument6 pagesHuman Sensesvicky7No ratings yet
- Visual DisordersDocument21 pagesVisual DisordersJobelle Acena100% (1)
- Oculomotor SystemDocument7 pagesOculomotor SystemZing WuNo ratings yet
- Oculomotor NerveDocument4 pagesOculomotor NerveHuzaifah IbrahimNo ratings yet
- ANATO - Campos VisuaisDocument11 pagesANATO - Campos VisuaishelbertpalmieroNo ratings yet
- Physiology of The Eye: Khorrami PH.DDocument119 pagesPhysiology of The Eye: Khorrami PH.Dkhorrami4No ratings yet
- subtitle-4Document2 pagessubtitle-4Lucas CruzNo ratings yet
- Physiology of EyeDocument17 pagesPhysiology of EyeHanna ShibuNo ratings yet
- Human EyeDocument12 pagesHuman Eyeanu rettiNo ratings yet
- Muscles of The EyeDocument3 pagesMuscles of The EyeAnna GarbiNo ratings yet
- CerebellumDocument26 pagesCerebellummaheswaransundaramoorthyNo ratings yet
- EyesDocument30 pagesEyesAlfredJoshuaMirandaNo ratings yet
- CHAPTER17 Anatomy and Physiology of OcularMotor SystemsDocument78 pagesCHAPTER17 Anatomy and Physiology of OcularMotor Systemsbenjumear_719607598No ratings yet
- Perception WordDocument3 pagesPerception WordNavya VaddimukkalaNo ratings yet
- Physiology of Eye. Physiology of VisionDocument27 pagesPhysiology of Eye. Physiology of VisionSmartcool So100% (1)
- Eye Movements Clinical CorrelationDocument8 pagesEye Movements Clinical CorrelationAsena TuiketeiNo ratings yet
- WPR 2Document5 pagesWPR 2ishika mohanNo ratings yet
- Human EyeDocument54 pagesHuman EyeRadu VisanNo ratings yet
- Dr. Herman - Physiology of EquilibriumDocument70 pagesDr. Herman - Physiology of EquilibriumAndrew Sabastian Geraldyno PaagoNo ratings yet
- Modelo Movimiento OjoDocument12 pagesModelo Movimiento OjoRafael EduardoNo ratings yet
- Handout (Sample)Document22 pagesHandout (Sample)judssalangsangNo ratings yet
- 4.3 Types of Eye MeovementDocument9 pages4.3 Types of Eye MeovementkksNo ratings yet
- Anatomy and Function of The EyeDocument3 pagesAnatomy and Function of The EyeAljhon DelfinNo ratings yet
- Eye Lecture GuideDocument46 pagesEye Lecture Guidemaj0% (1)
- 232 Coordinated Eye MovementsDocument2 pages232 Coordinated Eye MovementsVasilii LentitchiiNo ratings yet
- Biology - B9 - Summary 1 - IGCSEDocument16 pagesBiology - B9 - Summary 1 - IGCSE《Akari - San》 AMV ; 3No ratings yet
- Info Sheet 4 - 1Document8 pagesInfo Sheet 4 - 1Francis ObmergaNo ratings yet
- Anatomia y Fisiologuia General Libro-651-700Document50 pagesAnatomia y Fisiologuia General Libro-651-700Felipe Aguirre CórdovaNo ratings yet
- CSS Everyday Science Notes PDFDocument50 pagesCSS Everyday Science Notes PDFWaqas Gul100% (1)
- Co-Ordination and ResponseDocument19 pagesCo-Ordination and ResponseMuhammad ShahswarNo ratings yet
- Alde Annieska Learning Journal 5Document5 pagesAlde Annieska Learning Journal 5Maria AnnieskaNo ratings yet
- Eye 1989126Document17 pagesEye 1989126San Phạm ĐìnhNo ratings yet
- OphthalmologyDocument10 pagesOphthalmologyMohammad Farooq KhanNo ratings yet
- Visual-Vestibular Interaction - Basic Science To Clinical RelevanceDocument13 pagesVisual-Vestibular Interaction - Basic Science To Clinical RelevanceEnrico ArmatoNo ratings yet
- ACTIVITY 5 Special Senses FINALDocument2 pagesACTIVITY 5 Special Senses FINALJARISSA DIZON GALLAZANo ratings yet
- III, IV, VI Pair of Cranial NervesDocument5 pagesIII, IV, VI Pair of Cranial Nervesjalan_zNo ratings yet
- Sensing and Perceiving 4.2Document18 pagesSensing and Perceiving 4.2Dami TepedeNo ratings yet
- Cerebellum in Overall Motor ControlDocument5 pagesCerebellum in Overall Motor Controldncm.alejandroNo ratings yet
- OCULOPATHY - Disproves the orthodox and theoretical bases upon which glasses are so freely prescribed, and puts forward natural remedial methods of treatment for what are sometimes termed incurable visual defectsFrom EverandOCULOPATHY - Disproves the orthodox and theoretical bases upon which glasses are so freely prescribed, and puts forward natural remedial methods of treatment for what are sometimes termed incurable visual defectsNo ratings yet
- Visual Perception: Insights into Computational Visual ProcessingFrom EverandVisual Perception: Insights into Computational Visual ProcessingNo ratings yet
- Low Vision: Assessment and Educational Needs: A Guide to Teachers and ParentsFrom EverandLow Vision: Assessment and Educational Needs: A Guide to Teachers and ParentsNo ratings yet
- A Simple Guide to the Eye and Its Disorders, Diagnosis, Treatment and Related ConditionsFrom EverandA Simple Guide to the Eye and Its Disorders, Diagnosis, Treatment and Related ConditionsNo ratings yet
- Clinical Imaging: Small-Bowel Obstruction: State-of-the-Art Imaging and Its Role in Clinical ManagementDocument1 pageClinical Imaging: Small-Bowel Obstruction: State-of-the-Art Imaging and Its Role in Clinical ManagementDavis KallanNo ratings yet
- About NSD: On 28 February 1928, DR C.V Raman Had Announced The Discovery of TheDocument1 pageAbout NSD: On 28 February 1928, DR C.V Raman Had Announced The Discovery of TheDavis KallanNo ratings yet
- 42 Diabetic Retinopathy Classification and Typical LesionsDocument1 page42 Diabetic Retinopathy Classification and Typical LesionsDavis KallanNo ratings yet
- Radiology Is The Science That UsesDocument1 pageRadiology Is The Science That UsesDavis KallanNo ratings yet
- TamilDocument2 pagesTamilDavis KallanNo ratings yet
- Hindi Cinema (Bollywood)Document3 pagesHindi Cinema (Bollywood)Davis KallanNo ratings yet
- Marana Simhasanam Vanaprastham: Malayalam Cinema Is TheDocument1 pageMarana Simhasanam Vanaprastham: Malayalam Cinema Is TheDavis KallanNo ratings yet
- GQ (Indian Edition) : From Wikipedia, The Free EncyclopediaDocument2 pagesGQ (Indian Edition) : From Wikipedia, The Free EncyclopediaDavis KallanNo ratings yet
- Management Studies, Now Provides Its Mite in The Field of Higher Education Imparting The MostDocument1 pageManagement Studies, Now Provides Its Mite in The Field of Higher Education Imparting The MostDavis KallanNo ratings yet
- Tonometry PDFDocument1 pageTonometry PDFDavis KallanNo ratings yet
- Renal Concept MapDocument8 pagesRenal Concept MapDavis KallanNo ratings yet
- Renal Concept MapDocument8 pagesRenal Concept MapDavis KallanNo ratings yet
- Profile 14157Document3 pagesProfile 14157Davis KallanNo ratings yet
- Antimicrobials PDFDocument1 pageAntimicrobials PDFDavis KallanNo ratings yet
- Profile 14157Document3 pagesProfile 14157Davis KallanNo ratings yet
- Sexually Transmitted Infections PPT Presentation3749Document53 pagesSexually Transmitted Infections PPT Presentation3749Davis Kallan100% (3)
- Antivirals: Stages in Viral InfectionDocument1 pageAntivirals: Stages in Viral InfectionDavis KallanNo ratings yet
- Retinopathy of Prematurity - Leukocoria - Examining Neonates - Amblyopia - Squint Strabismus - (Not Covered Here in Detail Links)Document15 pagesRetinopathy of Prematurity - Leukocoria - Examining Neonates - Amblyopia - Squint Strabismus - (Not Covered Here in Detail Links)Davis KallanNo ratings yet
- Quick Review - Fossa of RosenmullerDocument4 pagesQuick Review - Fossa of RosenmullerDavis KallanNo ratings yet
- Log - 2022 8 8Document578 pagesLog - 2022 8 8Pedro Henrique Ferreira PrestjordNo ratings yet
- 310,534,414 Bahan Kimia Cair: Kode Uraian Nilai S/D 31 DESEMBER 2019 Jumlah RupiahDocument24 pages310,534,414 Bahan Kimia Cair: Kode Uraian Nilai S/D 31 DESEMBER 2019 Jumlah RupiahTri HandoyoNo ratings yet
- Initial Patient AssessmentDocument13 pagesInitial Patient AssessmentJolly RancherNo ratings yet
- Retinal Burn and Flash Blindness Data For Nuclear TestsDocument60 pagesRetinal Burn and Flash Blindness Data For Nuclear TestsNige CookNo ratings yet
- CA34a ReRevisedDocument9 pagesCA34a ReRevisedSS Malik100% (1)
- Daftar PustakaDocument2 pagesDaftar Pustakamelita Aditya SariNo ratings yet
- Anatomy of ConjuctivaDocument13 pagesAnatomy of ConjuctivaFaiz MazlanNo ratings yet
- Refractive ErrorsDocument73 pagesRefractive ErrorsAnggunBW100% (1)
- Pathway Psoriasis 1Document1 pagePathway Psoriasis 1Yani Indri YaniNo ratings yet
- ANATOMY and PHYSIOLOGY of CHRONIC OTITIS MEDIA-GCS Sa MED - ANNEX-maam ValezaDocument36 pagesANATOMY and PHYSIOLOGY of CHRONIC OTITIS MEDIA-GCS Sa MED - ANNEX-maam ValezaBonieve Pitogo NoblezadaNo ratings yet
- JI TicklerDocument25 pagesJI TicklerPatricia Anne ReyesNo ratings yet
- Structure and Function of The Human EyeDocument2 pagesStructure and Function of The Human EyefafafaNo ratings yet
- Structure and Function of EyesDocument11 pagesStructure and Function of EyesSamar Ameen SialNo ratings yet
- The Ear 2Document18 pagesThe Ear 2Aunties Hyd Kka DeewanaNo ratings yet
- Medical Department OpthomticDocument2 pagesMedical Department OpthomticdkhatriNo ratings yet
- Lens DislocationDocument13 pagesLens DislocationMuthia Fydiah JayantiNo ratings yet
- A Seminar Presentation On Special Senses Ana 803 (Neuroanatomy)Document24 pagesA Seminar Presentation On Special Senses Ana 803 (Neuroanatomy)Kolawole Adesina KehindeNo ratings yet
- Ocular Blunt Trauma: Loss of Sight From An Ice Hockey InjuryDocument2 pagesOcular Blunt Trauma: Loss of Sight From An Ice Hockey InjuryShirakawa AlmiraNo ratings yet
- Draize Irritation Test PDFDocument27 pagesDraize Irritation Test PDFhoythereNo ratings yet
- Anatomy and Physiology of The EarDocument2 pagesAnatomy and Physiology of The EarGunel AliyevaNo ratings yet
- Here Is Why Skincare Is ImportantDocument3 pagesHere Is Why Skincare Is ImportantKevin OkunguNo ratings yet
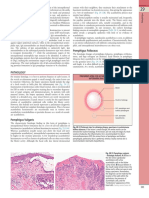
- Pemphigus Foliaceus: Fig. 29.13BDocument1 pagePemphigus Foliaceus: Fig. 29.13BCristian QuitoNo ratings yet
- Sensory Organs: EYE EAR Tongue Skin NoseDocument101 pagesSensory Organs: EYE EAR Tongue Skin NoseSaba PathyNo ratings yet
- ICSE Skin WorksheetDocument1 pageICSE Skin WorksheetpreetaNo ratings yet
- Eyelids, Conjunctiva, Cornea and Sclera: McquestionsDocument52 pagesEyelids, Conjunctiva, Cornea and Sclera: McquestionsHawwi Ismail100% (2)
- SightMD Referral Form PDFDocument2 pagesSightMD Referral Form PDFHENRYNo ratings yet
- NCM104 MS2 - Course Unit 13 - Review of The Anatomy and Physiology of The Eyes & AssessmentDocument5 pagesNCM104 MS2 - Course Unit 13 - Review of The Anatomy and Physiology of The Eyes & AssessmentJordz PlaciNo ratings yet
- Physiology of The Special SensesDocument31 pagesPhysiology of The Special SensesDemuel Dee L. BertoNo ratings yet
- I Look, I See: Learning ObjectivesDocument3 pagesI Look, I See: Learning ObjectivesRhex CalebNo ratings yet