Download as docx, pdf, or txt
You might also like
- VDA 6.3 - Jan 2023Document108 pagesVDA 6.3 - Jan 2023Cem Alsat95% (100)
- UndergroundG StyleHandgunMainManual PDFDocument46 pagesUndergroundG StyleHandgunMainManual PDFRick Gaines100% (2)
- Sandal Magna Primary SchoolDocument36 pagesSandal Magna Primary SchoolsaurabhNo ratings yet
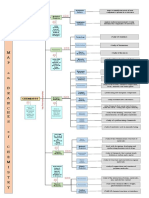
- Concept MapDocument1 pageConcept MapLesley Joy T. BaldonadoNo ratings yet
- MicropropagationDocument6 pagesMicropropagationEmmanuel OcanseyNo ratings yet
- In Vitro Plant Propagation - A Review - Kumar 2011Document12 pagesIn Vitro Plant Propagation - A Review - Kumar 2011Santiago RojasNo ratings yet
- Cell and Tissue Culture TechnologyDocument10 pagesCell and Tissue Culture TechnologyLegends FunanzaNo ratings yet
- Exercise 9 DevbiolabDocument4 pagesExercise 9 DevbiolabPrezyNo ratings yet
- Plant Tissue Culture: A Promising Tool of Quality Material Production With Special Reference To Micropropagation of BananaDocument26 pagesPlant Tissue Culture: A Promising Tool of Quality Material Production With Special Reference To Micropropagation of BananaFrance MalonzoNo ratings yet
- Biology Paper Plant Tissue Culture Aulia Putri KusumaDocument9 pagesBiology Paper Plant Tissue Culture Aulia Putri KusumaAulia Putri KusumaNo ratings yet
- Kumar-2011-In Vitro Plant Propagation A ReviewDocument13 pagesKumar-2011-In Vitro Plant Propagation A ReviewJuanmanuelNo ratings yet
- Application of Tissue Culture in Plant BreedingDocument7 pagesApplication of Tissue Culture in Plant BreedingNasir Hussain Faraz89% (9)
- Plant Tissue Culture: Biology-100 Section: 7 DR - Jebunnahar KhandakarDocument17 pagesPlant Tissue Culture: Biology-100 Section: 7 DR - Jebunnahar Khandakarijaj imran100% (2)
- send Adrae Anderson BIOT3113 Lab 2 Tissue CultureDocument17 pagessend Adrae Anderson BIOT3113 Lab 2 Tissue Culturetashy richardsNo ratings yet
- Biotech BookDocument31 pagesBiotech BookMehedi HossainNo ratings yet
- Priyadarshani P. Mohapatra and V.K. BatraDocument7 pagesPriyadarshani P. Mohapatra and V.K. BatraAndreaMolinaDuránNo ratings yet
- Elaboration of Micropropagation Protocol For Vaccinium Corymbosum CV Sunt Blue GiantDocument12 pagesElaboration of Micropropagation Protocol For Vaccinium Corymbosum CV Sunt Blue GiantPriscila GuiselleNo ratings yet
- Plant Tissue CultureDocument7 pagesPlant Tissue CultureAllwin SamNo ratings yet
- Importance of Tissue Culture in AgricultureDocument4 pagesImportance of Tissue Culture in AgricultureTaylor Isaac100% (1)
- Chapter 2 (Animal Cell Culture, Micropropagation, IVF, Cloning)Document88 pagesChapter 2 (Animal Cell Culture, Micropropagation, IVF, Cloning)Batul KhuzemaNo ratings yet
- Plant Tissue CultureDocument3 pagesPlant Tissue CulturepradeepNo ratings yet
- Plant Tissue CultureDocument5 pagesPlant Tissue Culturesakshisangwan02No ratings yet
- 15240338946QuadrantIE TEXTDocument15 pages15240338946QuadrantIE TEXTSelvakapoorNo ratings yet
- Disadvantages: Somaclonal Variation Is The Variation Seen in Plants That Have Been Produced by Plant TissueDocument4 pagesDisadvantages: Somaclonal Variation Is The Variation Seen in Plants That Have Been Produced by Plant TissueEN1036Vivek SinghNo ratings yet
- Tissue Culture in PlantsDocument83 pagesTissue Culture in PlantsEN1036Vivek Singh100% (1)
- Micro Propagation: Some of The Important Plants That Have Been Micro Propagated On Large Scales AreDocument59 pagesMicro Propagation: Some of The Important Plants That Have Been Micro Propagated On Large Scales AreVijayNo ratings yet
- General Techniques of Plant Tissue Culture: November 2015Document31 pagesGeneral Techniques of Plant Tissue Culture: November 2015Botany DepartmentNo ratings yet
- Tissue CultureDocument15 pagesTissue Culturebudharam choudharyNo ratings yet
- Application of Plant Tisse CultureDocument23 pagesApplication of Plant Tisse CultureSujalNo ratings yet
- Plant Tissue CultureDocument26 pagesPlant Tissue CultureNAVEENANo ratings yet
- Ijair 3275 FinalDocument8 pagesIjair 3275 Finaloztrk.meltmNo ratings yet
- History of Plant Tissue CultureDocument14 pagesHistory of Plant Tissue CultureSTAR PRINTINGNo ratings yet
- Introduction To Agriculture Biotechnology and Its ApplicationsDocument5 pagesIntroduction To Agriculture Biotechnology and Its ApplicationsZohaZehraNo ratings yet
- Somaclonal Variation in Tissue Culture: A Case Study With OliveDocument28 pagesSomaclonal Variation in Tissue Culture: A Case Study With Olivekj185No ratings yet
- Large Scale Production Hairy RootDocument54 pagesLarge Scale Production Hairy RootMukund ShankarNo ratings yet
- Plant Tissue Culture - Biotechnology - Online Microbiology NotesDocument5 pagesPlant Tissue Culture - Biotechnology - Online Microbiology NotesBaikuntha SabarNo ratings yet
- Jyoti Singh and Anuj KumarDocument11 pagesJyoti Singh and Anuj Kumarudayakundu143No ratings yet
- Plant Tissue CultureDocument9 pagesPlant Tissue CulturedushyantrockyNo ratings yet
- ThiartDocument7 pagesThiartmanojbanNo ratings yet
- Microbial Growth - An Overview ScienceDirect TopicsDocument1 pageMicrobial Growth - An Overview ScienceDirect Topicssh.dabir1970No ratings yet
- Tissue Culture As A Plant Production Technique For Horticultural CropsDocument8 pagesTissue Culture As A Plant Production Technique For Horticultural CropsSoummyadip RoyNo ratings yet
- Pl. Tissue Culture 3Document23 pagesPl. Tissue Culture 3Akash RoyNo ratings yet
- Plants: Cellular, Molecular, and Physiological Aspects of in Vitro Plant RegenerationDocument20 pagesPlants: Cellular, Molecular, and Physiological Aspects of in Vitro Plant RegenerationPham Thi My TramNo ratings yet
- Practocal Manual On Plant Tissue CultureDocument130 pagesPractocal Manual On Plant Tissue CultureĐương Anh CaoNo ratings yet
- Thesis On Micropropagation of BananaDocument4 pagesThesis On Micropropagation of BananaMary Calkins100% (2)
- Tissue Culture ThesisDocument5 pagesTissue Culture ThesisBestPaperWritersKansasCity100% (2)
- Mass Multiplication Procedure For Tissue CultureDocument28 pagesMass Multiplication Procedure For Tissue CultureDeepak SharmaNo ratings yet
- Plant Tissue Culture PHD ThesisDocument6 pagesPlant Tissue Culture PHD Thesisemilycobbinsdesmoines100% (2)
- Plant Tissue Cultur1Document74 pagesPlant Tissue Cultur1Amit SinghNo ratings yet
- Proposal KentangDocument12 pagesProposal Kentangrico100% (1)
- 760D84F31753Document5 pages760D84F31753Spam SpamNo ratings yet
- Plant BiotechnologyDocument7 pagesPlant BiotechnologyJhun Lerry TayanNo ratings yet
- Plant Tissue Culture A Promising Tool of Quality MaterialDocument27 pagesPlant Tissue Culture A Promising Tool of Quality MaterialkusumawardatiNo ratings yet
- Plant Tissue Culture-IntroductionDocument11 pagesPlant Tissue Culture-Introductionnehabano336No ratings yet
- Term Paper On: Transgenic PlantsDocument14 pagesTerm Paper On: Transgenic PlantsVikal RajputNo ratings yet
- PTC QaDocument71 pagesPTC Qanavid akibNo ratings yet
- From Micropropagation To MicroponicsDocument5 pagesFrom Micropropagation To Microponicsbrijmohansingh401124No ratings yet
- Micropropagation of Banana ThesisDocument7 pagesMicropropagation of Banana ThesisWhereCanIFindSomeoneToWriteMyPaperCanada100% (2)
- Industrial MicroorganismsDocument7 pagesIndustrial MicroorganismsriddhisnewworldNo ratings yet
- Theory: Micropropagation Plant TissueDocument17 pagesTheory: Micropropagation Plant TissuekzrdurNo ratings yet
- Somatic Embryogenesis for Micropropagation of Coconut (Cocos nucifera L.)From EverandSomatic Embryogenesis for Micropropagation of Coconut (Cocos nucifera L.)No ratings yet
- Outlines of dairy bacteriology A concise manual for the use of students in dairyingFrom EverandOutlines of dairy bacteriology A concise manual for the use of students in dairyingNo ratings yet
- Outlines of Dairy Bacteriology, 8th edition A Concise Manual for the Use of Students in DairyingFrom EverandOutlines of Dairy Bacteriology, 8th edition A Concise Manual for the Use of Students in DairyingNo ratings yet
- ISquare - Setup Guide For Fresh UsersDocument12 pagesISquare - Setup Guide For Fresh UsersWQDNo ratings yet
- The Wholly BibleDocument10 pagesThe Wholly BibleFred_Carpenter_12No ratings yet
- Campaign For Tobacco Free KidsDocument1 pageCampaign For Tobacco Free KidsNurses For Tobacco ControlNo ratings yet
- Expect Respect ProgramDocument2 pagesExpect Respect ProgramCierra Olivia Thomas-WilliamsNo ratings yet
- Yohe Nitsumi: GH 15.00 GH 15.00 GH 15.00Document3 pagesYohe Nitsumi: GH 15.00 GH 15.00 GH 15.00lYNX GHNo ratings yet
- Draft Local Budget Circular Re Grant of Honorarium To SK Officials - v1Document4 pagesDraft Local Budget Circular Re Grant of Honorarium To SK Officials - v1rodysseusNo ratings yet
- Sample Thesis 2Document127 pagesSample Thesis 2Carlo Troy AcelottNo ratings yet
- Break Even AnalysisDocument18 pagesBreak Even AnalysisSMHE100% (10)
- Full Download Art of Leadership 5th Edition Manning Test Bank PDF Full ChapterDocument36 pagesFull Download Art of Leadership 5th Edition Manning Test Bank PDF Full Chaptermasqueedenized8a43l100% (19)
- ICF Team Coaching Competencies: Moving Beyond One-to-One CoachingDocument21 pagesICF Team Coaching Competencies: Moving Beyond One-to-One CoachingMarko PyNo ratings yet
- General: Specific: Leading Statement:: TITLE: A Healthy LifestyleDocument2 pagesGeneral: Specific: Leading Statement:: TITLE: A Healthy LifestyleM Arshad M AmirNo ratings yet
- Butterflies and MothsDocument59 pagesButterflies and MothsReegen Tee100% (3)
- Assignment-II UNIT-I Topic: AveragesDocument3 pagesAssignment-II UNIT-I Topic: AveragesAjay JaginiNo ratings yet
- December 2023 1Document14 pagesDecember 2023 1febri.putra7280No ratings yet
- Edward SapirDocument10 pagesEdward Sapirparand2003No ratings yet
- Petitioner vs. VS.: Third DivisionDocument12 pagesPetitioner vs. VS.: Third DivisionAnonymous QR87KCVteNo ratings yet
- SY BSC IT SEM IV CORE JAVA UNIT 1Document56 pagesSY BSC IT SEM IV CORE JAVA UNIT 1stellaNo ratings yet
- Adavantages of Historical ResearchDocument2 pagesAdavantages of Historical ResearchClemence JeniferNo ratings yet
- Karakteristik Material Baja St.37 Dengan Temperatur Dan Waktu Pada Uji Heat Treatment Menggunakan FurnaceDocument14 pagesKarakteristik Material Baja St.37 Dengan Temperatur Dan Waktu Pada Uji Heat Treatment Menggunakan FurnaceYudii NggiNo ratings yet
- Kinds of Sentences and Their Punctuation: Coordinating Conjunction Conjunctive Adverb SemicolonDocument41 pagesKinds of Sentences and Their Punctuation: Coordinating Conjunction Conjunctive Adverb SemicolonMarwaNo ratings yet
- Alternative Substrates As A Native Oyster (Crassostrea Virginica) Reef Restoration Strategy in Chesapeake BayDocument286 pagesAlternative Substrates As A Native Oyster (Crassostrea Virginica) Reef Restoration Strategy in Chesapeake BayMandi MynyddNo ratings yet
- Investments: Analysis and Behavior: Chapter 1-IntroductionDocument26 pagesInvestments: Analysis and Behavior: Chapter 1-IntroductionJeralyn MacarealNo ratings yet
- Ahad NaamahDocument2 pagesAhad NaamahedoolawNo ratings yet
- Ncert Solutions Class 6 Civics Chapter 3 PDFDocument2 pagesNcert Solutions Class 6 Civics Chapter 3 PDFSabaNo ratings yet
- World Red Cross DayDocument17 pagesWorld Red Cross DayAshwani K SharmaNo ratings yet
- Comparison of Alternatives NotesDocument10 pagesComparison of Alternatives NotesLibyaFlowerNo ratings yet