Download as pdf or txt
You might also like
- Activation of CD8 T Cells Induces Expression of CD4, Which Functions As A Chemotactic ReceptorDocument6 pagesActivation of CD8 T Cells Induces Expression of CD4, Which Functions As A Chemotactic ReceptorKevin MaiseyNo ratings yet
- Dejaco-Et-Al 06 - (Imbalance of Regulatory T Cells in Human Autoimmune Diseases)Document12 pagesDejaco-Et-Al 06 - (Imbalance of Regulatory T Cells in Human Autoimmune Diseases)Luana DiEmmeNo ratings yet
- Ek 2006Document6 pagesEk 2006ᄋᄋNo ratings yet
- Chloremphenicol Leukemia LinkedDocument17 pagesChloremphenicol Leukemia LinkedpadbidriNo ratings yet
- CTLA-4 (CD152) Controls Homeostasis and Suppressive Capacity of Regulatory T Cells in MiceDocument10 pagesCTLA-4 (CD152) Controls Homeostasis and Suppressive Capacity of Regulatory T Cells in MiceMaxime PorcoNo ratings yet
- 2022 Article 210Document13 pages2022 Article 210uttamNo ratings yet
- Prolactin Increase Frecuency of T Helper FolicularDocument15 pagesProlactin Increase Frecuency of T Helper Folicularauroragamez2015No ratings yet
- Abstracts For The 27th Annual Scientific Meeting of The Society For Immunotherapy of Cancer (SITC) PDFDocument71 pagesAbstracts For The 27th Annual Scientific Meeting of The Society For Immunotherapy of Cancer (SITC) PDFhigginscribdNo ratings yet
- CD28AJPDocument8 pagesCD28AJPAndrás KomócsiNo ratings yet
- His 12031Document4 pagesHis 12031JeevikaGoyalNo ratings yet
- A Systematic Approach To Diagnosis of Mature T-Cell LeukemiasDocument9 pagesA Systematic Approach To Diagnosis of Mature T-Cell LeukemiasRiki SanNo ratings yet
- Aggressive T - Cell Lymphomas 2021 UpdatesDocument41 pagesAggressive T - Cell Lymphomas 2021 UpdatesAnnaNo ratings yet
- Tumour Grade Significantly Correlates With Total Dysfunction of Tumour Tissue-Infiltrating Lymphocytes in Renal Cell CarcinomaDocument13 pagesTumour Grade Significantly Correlates With Total Dysfunction of Tumour Tissue-Infiltrating Lymphocytes in Renal Cell Carcinomasara.madkour99No ratings yet
- Anumanthan J Immunol-1998Document12 pagesAnumanthan J Immunol-1998raje bakshaNo ratings yet
- 1 s2.0 S0145305X1100303X MainDocument13 pages1 s2.0 S0145305X1100303X MainEdu AlmeidaNo ratings yet
- STAT3 Regulates Cytokine-Mediated Generation of Inflammatory Helper T CellsDocument6 pagesSTAT3 Regulates Cytokine-Mediated Generation of Inflammatory Helper T Cellssa571922No ratings yet
- Spittler Et Al. - 1997 - A Glutamine Deficiency Impairs The Function of CulDocument3 pagesSpittler Et Al. - 1997 - A Glutamine Deficiency Impairs The Function of CulMATHILDE MAGRONo ratings yet
- Gynecologic Oncology: John R. Webb, Katy Milne, David R. Kroeger, Brad H. NelsonDocument10 pagesGynecologic Oncology: John R. Webb, Katy Milne, David R. Kroeger, Brad H. NelsonAnonymous ilIW1YgNo ratings yet
- Abstract NKI KnoopsDocument1 pageAbstract NKI Knoopsakbar_rozaaqNo ratings yet
- Immune Response After Autologous Hematopoietic Stem Cell Transplantation in Type 1 Diabetes MellitusDocument10 pagesImmune Response After Autologous Hematopoietic Stem Cell Transplantation in Type 1 Diabetes Mellitusaini nazilaNo ratings yet
- Original Article: Abnormal Humans WithDocument11 pagesOriginal Article: Abnormal Humans WithArie kusuma WijayaNo ratings yet
- MHC Class and Class Product by Malignant T Cells: Relationships CD1a, and F2-MicroglobulinDocument5 pagesMHC Class and Class Product by Malignant T Cells: Relationships CD1a, and F2-MicroglobulindavdavdavdavdavdavdaNo ratings yet
- 5TOAUTOJDocument5 pages5TOAUTOJRika FitriaNo ratings yet
- Alter 2004Document8 pagesAlter 2004Ivan VazquezNo ratings yet
- Rheumatology-2014-Liu-rheumatology-keu316 HormonaDocument11 pagesRheumatology-2014-Liu-rheumatology-keu316 HormonaBluna0291No ratings yet
- Dendritic Cells and Humoral Immunity in Humans: ReviewDocument5 pagesDendritic Cells and Humoral Immunity in Humans: ReviewCarlos GuerreroNo ratings yet
- CD107 NK Cell 2004Document8 pagesCD107 NK Cell 2004Lu LizNo ratings yet
- Cell DiabetesDocument8 pagesCell DiabetesSusana Calderón ToledoNo ratings yet
- Immunocytochemical Studies of Cyclin D1 and P16 INK4a in Acute Lymphoblastic Leukemic PatientsDocument11 pagesImmunocytochemical Studies of Cyclin D1 and P16 INK4a in Acute Lymphoblastic Leukemic PatientsLotfy MaktaryNo ratings yet
- 2012 MC Proteomics - Proteomics and Transcriptomics Approach To Identify Leukemic Stem Cell MarkersDocument34 pages2012 MC Proteomics - Proteomics and Transcriptomics Approach To Identify Leukemic Stem Cell MarkersJosé Nivaldo Ifatunmibi Aworeni OdusolaNo ratings yet
- CLL and ERs Manuscript.Document15 pagesCLL and ERs Manuscript.Nadeem khanNo ratings yet
- Citosol (Thiamylal Sodium) Triggers Apoptosis and Affects Gene Expressions of Murine Leukemia RAW 264.7 Cells - RS-C Wu, C-S Yu, 2Document1 pageCitosol (Thiamylal Sodium) Triggers Apoptosis and Affects Gene Expressions of Murine Leukemia RAW 264.7 Cells - RS-C Wu, C-S Yu, 2Alondra MaldonadoNo ratings yet
- The Role of Natural Killer Cells in Chronic Myeloid LeukemiaDocument5 pagesThe Role of Natural Killer Cells in Chronic Myeloid LeukemiaRicardo MeloNo ratings yet
- CD8+CD282 Cells and CD4+CD25+ Regulatory T Cells in The Peripheral Blood of Advanced Stage Lung Cancer PatientsDocument6 pagesCD8+CD282 Cells and CD4+CD25+ Regulatory T Cells in The Peripheral Blood of Advanced Stage Lung Cancer PatientsEN Ka ERNo ratings yet
- Min Dong, Guozhen Zhang, Jie Meng, Biou Liu, Duanfeng Jiang, and Feng LiuDocument22 pagesMin Dong, Guozhen Zhang, Jie Meng, Biou Liu, Duanfeng Jiang, and Feng LiuEtienne UiSNo ratings yet
- Innate Immunity 3.1 - Natural Killer Cells (NK) and Natural Killer T (NKT) Cells in PEDocument13 pagesInnate Immunity 3.1 - Natural Killer Cells (NK) and Natural Killer T (NKT) Cells in PEShali NovizarNo ratings yet
- Stem CellsDocument12 pagesStem CellsLunaNo ratings yet
- The Genetic Background of Pancreatic Cancer - Genes That Might Be Biomarkers or Indicators of Metastasis To The Lung.Document1 pageThe Genetic Background of Pancreatic Cancer - Genes That Might Be Biomarkers or Indicators of Metastasis To The Lung.Laur NeyNo ratings yet
- How I Treat T-Cell Acute Lymphoblastic Leukemia in AdultsDocument10 pagesHow I Treat T-Cell Acute Lymphoblastic Leukemia in AdultsIndahNo ratings yet
- 2010 ER Stress Induction in T CellDocument6 pages2010 ER Stress Induction in T CellRaul ReyesNo ratings yet
- Spittler Et Al. - 1995 - Influence of Glutamine On The Phenotype and FunctiDocument6 pagesSpittler Et Al. - 1995 - Influence of Glutamine On The Phenotype and FunctiMATHILDE MAGRONo ratings yet
- Ichikawa 1999Document4 pagesIchikawa 1999Araceli Enríquez OvandoNo ratings yet
- Thl/Th2 Profiles Tuberculosis, Proliferation Cytokine of Blood Lymphocytes Mycobacterial AntigensDocument6 pagesThl/Th2 Profiles Tuberculosis, Proliferation Cytokine of Blood Lymphocytes Mycobacterial AntigensAdolfo Arturo Ccencho VacasNo ratings yet
- C-Rel Is Crucial For Lymphocyte Proliferation But Dispensable For T Cell Effector FunctionDocument11 pagesC-Rel Is Crucial For Lymphocyte Proliferation But Dispensable For T Cell Effector FunctionFrontiersNo ratings yet
- Normal Value CytokinesDocument7 pagesNormal Value Cytokinesdiana.alyNo ratings yet
- Imunology PregnancyDocument9 pagesImunology PregnancyimeldafitriNo ratings yet
- PCK 9Document10 pagesPCK 9hendarto ArifNo ratings yet
- Rabbit Antithymocyte Globulin (Thymoglobulin®) Impairs The Thymic Out-Put of Both Conventional and Regulatory CD4 T Cells After Allogeneic Hematopoietic Stem Cell Transplantation in Adult PatientsDocument8 pagesRabbit Antithymocyte Globulin (Thymoglobulin®) Impairs The Thymic Out-Put of Both Conventional and Regulatory CD4 T Cells After Allogeneic Hematopoietic Stem Cell Transplantation in Adult PatientsGhanta Ranjith KumarNo ratings yet
- MCB00436-22-Merged - PDF MergedDocument47 pagesMCB00436-22-Merged - PDF MergedHector Ivan Saldivar CeronNo ratings yet
- Articol MYD88, PIM-1Document18 pagesArticol MYD88, PIM-1Ionut PoinareanuNo ratings yet
- Edwin Darmawan: Moderator: Dr. Dr. Hani Susianti, SP - PK (K)Document48 pagesEdwin Darmawan: Moderator: Dr. Dr. Hani Susianti, SP - PK (K)Edwin DarmawanNo ratings yet
- 2001 - Regulation of IFN-γ signaling is essential for the cytotoxic activity of CD8+Document10 pages2001 - Regulation of IFN-γ signaling is essential for the cytotoxic activity of CD8+顏世隆No ratings yet
- J Exp Med 1995 Krummel 459 65Document7 pagesJ Exp Med 1995 Krummel 459 65Fernanda RibeiroNo ratings yet
- Ijcem 1309001Document10 pagesIjcem 1309001Wulandari Sri RezekiNo ratings yet
- Benevides2018 PDFDocument15 pagesBenevides2018 PDFNayche TortatoNo ratings yet
- Increased Number of Tc17 and Correlation With Th17 Cells in Patients With Immune ThrombocytopeniaDocument7 pagesIncreased Number of Tc17 and Correlation With Th17 Cells in Patients With Immune Thrombocytopeniadia puspitaNo ratings yet
- CBM Final PDFDocument7 pagesCBM Final PDFSamah Abd ElhamidNo ratings yet
- Cancer Immunosurveillance: AngiogenesisDocument2 pagesCancer Immunosurveillance: AngiogenesisBrenda Bastos LevanoNo ratings yet
- Co-signal Molecules in T Cell Activation: Immune Regulation in Health and DiseaseFrom EverandCo-signal Molecules in T Cell Activation: Immune Regulation in Health and DiseaseMiyuki AzumaNo ratings yet
- Tumor Immune Microenvironment in Cancer Progression and Cancer TherapyFrom EverandTumor Immune Microenvironment in Cancer Progression and Cancer TherapyPawel KalinskiNo ratings yet
- Bowel TrainingDocument2 pagesBowel Trainingdickie.gdrNo ratings yet
- Program - Neurogastro 2019 PDFDocument14 pagesProgram - Neurogastro 2019 PDFloloasbNo ratings yet
- JaundiceDocument29 pagesJaundiceMurali TiarasanNo ratings yet
- Surgical Ear Diseases of The Dog - An OverviewDocument9 pagesSurgical Ear Diseases of The Dog - An OverviewPavel VokrouhlíkNo ratings yet
- Renal FailureDocument41 pagesRenal Failure12046No ratings yet
- Book ReviewDocument13 pagesBook ReviewMoh AzharNo ratings yet
- Best Zapper: Who Is Dr. Hulda Clark?Document10 pagesBest Zapper: Who Is Dr. Hulda Clark?DR. NAFEES IQBALNo ratings yet
- Conf Brochure Terpenois AromatherapyDocument8 pagesConf Brochure Terpenois Aromatherapyafr307No ratings yet
- PH2 DatasetDocument20 pagesPH2 DatasetPopa DariaNo ratings yet
- Tonation - Spectro-Chrome MetryDocument5 pagesTonation - Spectro-Chrome MetryitounosNo ratings yet
- Goris PDFDocument24 pagesGoris PDFAndrew WongNo ratings yet
- SIMDUT-whmis ENGLISHDocument1 pageSIMDUT-whmis ENGLISHwacwac2011No ratings yet
- Cucumber As A Face Toner For Acne: Julia Bawayan Dan Oligo Alexander Estigoy Joseph TuralbaDocument18 pagesCucumber As A Face Toner For Acne: Julia Bawayan Dan Oligo Alexander Estigoy Joseph Turalbastella bawayanNo ratings yet
- Radiation Therapy and You: Support For People With CancerDocument68 pagesRadiation Therapy and You: Support For People With Cancerrazvan_nNo ratings yet
- RhabdomyosarcomaDocument11 pagesRhabdomyosarcomaFrancia ToledanoNo ratings yet
- Pre Malignant and Malignant Lesions of Cervix: Gynaec Unit 5Document147 pagesPre Malignant and Malignant Lesions of Cervix: Gynaec Unit 5FarazNo ratings yet
- Lab 4 5 Renal Functions TestDocument6 pagesLab 4 5 Renal Functions TestMustafa MahdiNo ratings yet
- Ertuglifozin DM IIDocument2 pagesErtuglifozin DM IIMr. LNo ratings yet
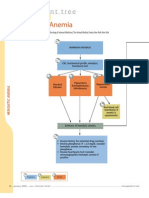
- Diagnostic Tree - Hemolytic AnemiaDocument2 pagesDiagnostic Tree - Hemolytic AnemiaЂорђеNo ratings yet
- Bahasa Inggris 3Document9 pagesBahasa Inggris 3vinavzenirNo ratings yet
- Problem Solving With AnswersDocument50 pagesProblem Solving With Answersapi-3814958No ratings yet
- Mammography Technique and TypesDocument3 pagesMammography Technique and Typesshuja behnaNo ratings yet
- 4258 FullDocument11 pages4258 FullshazaNo ratings yet
- You Can Quit Tobacco - by Vance FerrellDocument65 pagesYou Can Quit Tobacco - by Vance FerrellAdventist_Truth100% (1)
- TireotoxicozaDocument3 pagesTireotoxicozaDragos PopoviciNo ratings yet
- KJ - Ppt..chromosome DefectsDocument84 pagesKJ - Ppt..chromosome DefectsGerald CatiponNo ratings yet
- Ultrastructure and Functions of CementumDocument106 pagesUltrastructure and Functions of CementumJaya Shakthi0% (1)
- 2001 Self-Assessment Examination For Residents (Sae-R) Multiple-Choice Questions Booklet Abridged VersionDocument23 pages2001 Self-Assessment Examination For Residents (Sae-R) Multiple-Choice Questions Booklet Abridged VersionSoumabho DasNo ratings yet
- The Beginners Guide To Micro NichesDocument22 pagesThe Beginners Guide To Micro NicheselsaliliNo ratings yet
- Approach To Space Occupying LesionDocument14 pagesApproach To Space Occupying LesionFatima JamshaidNo ratings yet