
Chapter 10
Chapter 10
You might also like
- Life Orientation Term 3 Grade 11Document11 pagesLife Orientation Term 3 Grade 11farihah75% (4)
- Diabetic Nephropathy Case StudyDocument61 pagesDiabetic Nephropathy Case Studyoxidalaj98% (63)
- EvolutionDocument4 pagesEvolutionChaudhry AzeemNo ratings yet
- From Purgatorius Ceratops To Homo Sapiens: I. Primate Evolutionary HistoryDocument10 pagesFrom Purgatorius Ceratops To Homo Sapiens: I. Primate Evolutionary HistoryquiquebravoNo ratings yet
- SBB 6014 Evolution of MammalsDocument19 pagesSBB 6014 Evolution of MammalsJauha HanafiNo ratings yet
- Article Fossil AnimalsDocument4 pagesArticle Fossil AnimalsmozartNo ratings yet
- Mammal: MammalsDocument13 pagesMammal: Mammalsdaitya99No ratings yet
- Ars Technica - The Paja FormationDocument4 pagesArs Technica - The Paja Formationmilton74No ratings yet
- SynapsidsDocument7 pagesSynapsidsapi-407347220No ratings yet
- The Earliest Possible Hominids: Sahelanthropus TchadensisDocument7 pagesThe Earliest Possible Hominids: Sahelanthropus Tchadensisdod_nurNo ratings yet
- PDF Brusatteetal2010TyrannosaurPaleobiologyDocument24 pagesPDF Brusatteetal2010TyrannosaurPaleobiologyArnab DasNo ratings yet
- Evidence of Evolution On Animal VertebrateDocument9 pagesEvidence of Evolution On Animal Vertebratejohanes manaluNo ratings yet
- Septianisa UlfalahDocument12 pagesSeptianisa UlfalahSeptianisa UlfalahNo ratings yet
- Ucsp (Lesson 1)Document28 pagesUcsp (Lesson 1)united nationNo ratings yet
- Chapter 4Document172 pagesChapter 4Aan AkmalNo ratings yet
- UE Evograms Jaws To EarsDocument3 pagesUE Evograms Jaws To Earsskline3No ratings yet
- Dinosaur: Dinosaurs Are A Diverse Group of Animals of TheDocument5 pagesDinosaur: Dinosaurs Are A Diverse Group of Animals of ThebabublsNo ratings yet
- Dino Jaws Information SheetsDocument55 pagesDino Jaws Information SheetsJacob PruittNo ratings yet
- Evolution Epidemiology and Etiology of Temporomandibular Joint DisordersDocument6 pagesEvolution Epidemiology and Etiology of Temporomandibular Joint DisordersCM Panda CedeesNo ratings yet
- App 20100005Document22 pagesApp 20100005Ghaier KazmiNo ratings yet
- Life-History Evolution in ReptilesDocument11 pagesLife-History Evolution in ReptilesjeraldNo ratings yet
- Anthropological Concepts in Clinical OrthodonticsDocument42 pagesAnthropological Concepts in Clinical Orthodonticsayesha rathore100% (1)
- Freeman. 1981. Correspondence of Food Habits and Morphology in Insectivorous BatsDocument9 pagesFreeman. 1981. Correspondence of Food Habits and Morphology in Insectivorous BatsHenry CondoriNo ratings yet
- 216) Escanear0001Document1 page216) Escanear0001Beatriz MogollonNo ratings yet
- 216) Escanear0001Document1 page216) Escanear0001Beatriz MogollonNo ratings yet
- Bed 130 - Week 1 TaskDocument27 pagesBed 130 - Week 1 TaskPaul Ivan L. PazNo ratings yet
- Origins of CetaceansDocument17 pagesOrigins of CetaceansVengadesh Krishna KumarNo ratings yet
- The Origin and Evolution of DinosaursDocument55 pagesThe Origin and Evolution of DinosaursTiago Savignon Cardoso MachadoNo ratings yet
- Biodiversity and EvolutionDocument99 pagesBiodiversity and EvolutionMaria Jamilla R. Pua100% (1)
- AEB 317 Introductory EntomologyDocument35 pagesAEB 317 Introductory EntomologyOgualu FavourNo ratings yet
- Earth History - The Geologic Time ScaleDocument26 pagesEarth History - The Geologic Time ScaleCarminda Subaba CastilloNo ratings yet
- GEN-BIO-2-History of Life On EarthDocument22 pagesGEN-BIO-2-History of Life On EarthGeminiNo ratings yet
- Cope's Rule and The Adaptive Landscape of Dinosaur Body Size EvolutionDocument36 pagesCope's Rule and The Adaptive Landscape of Dinosaur Body Size Evolutionbraydenstoch777No ratings yet
- Etymology: DimetrodonDocument5 pagesEtymology: DimetrodonAjay SinghNo ratings yet
- Fossil Records PDFDocument10 pagesFossil Records PDFShane CubioNo ratings yet
- Paleogene and NeogeneDocument41 pagesPaleogene and Neogenefredcramer081No ratings yet
- Half A Billion Years of BackbonesDocument1 pageHalf A Billion Years of BackbonesM HanifNo ratings yet
- Homo HobbitDocument4 pagesHomo HobbitAndrew JonesNo ratings yet
- Fossils & EvolutionDocument11 pagesFossils & EvolutionDaniel BerryNo ratings yet
- Growth Patterns of Fossil Vertebrates As Deduced From Bone Microstructure: Case Studies From IndiaDocument12 pagesGrowth Patterns of Fossil Vertebrates As Deduced From Bone Microstructure: Case Studies From IndiaOMSURYACHANDRASIVANo ratings yet
- #2 Chapter IIDocument17 pages#2 Chapter IIAngelyn Bantilo SerronaNo ratings yet
- Sauropod Dinosaur FossilDocument3 pagesSauropod Dinosaur FossilWestern Ang'ong'aNo ratings yet
- Darwin's Theory EssayDocument6 pagesDarwin's Theory EssayLogkeetha SarckunadasanNo ratings yet
- Bernard ReviewerDocument2 pagesBernard ReviewerBernard BrowneNo ratings yet
- The Existence of TrilobitesDocument4 pagesThe Existence of Trilobitesvidur_talrejaNo ratings yet
- The Origin of Animal Body PlansDocument15 pagesThe Origin of Animal Body PlansDiego LoboNo ratings yet
- Diet and The Evolution of The Earliest Human Ancestors: Mark F. Teaford and Peter S. UngarDocument6 pagesDiet and The Evolution of The Earliest Human Ancestors: Mark F. Teaford and Peter S. UngarRaul Diaz TorresNo ratings yet
- Rocio Huizar Instructor Teri Potter Anthropology 1020-012Document5 pagesRocio Huizar Instructor Teri Potter Anthropology 1020-012api-311470705No ratings yet
- Macroevolution and The Formation of New SpeciesDocument19 pagesMacroevolution and The Formation of New Speciesmichelle ann sabilonaNo ratings yet
- Haas 1998Document33 pagesHaas 1998lassaad.el.hanafiNo ratings yet
- UNIT 1palaeontology Handout..Document8 pagesUNIT 1palaeontology Handout..getnet gizachewNo ratings yet
- Lesson 5.1 SummaryDocument4 pagesLesson 5.1 SummaryJaime MallaNo ratings yet
- Overview of ProjectDocument14 pagesOverview of Projectofmephotos4No ratings yet
- Population Structure of The Gomphothere Stegomastodon Waringi (Mammalia: Proboscidea: Gomphotheriidae) From The Pleistocene of BrazilDocument14 pagesPopulation Structure of The Gomphothere Stegomastodon Waringi (Mammalia: Proboscidea: Gomphotheriidae) From The Pleistocene of BrazilKanekoNo ratings yet
- Blueprint of LifeDocument11 pagesBlueprint of Lifetimeflies23No ratings yet
- General Description: Triceratops Megalosaurus IguanodonDocument31 pagesGeneral Description: Triceratops Megalosaurus IguanodonRicardo Daniel Aranda ApolinarioNo ratings yet
- 4-Rodriguez-De La Rosa - Etal - 2021 - Fish-trace-fossil-KDocument10 pages4-Rodriguez-De La Rosa - Etal - 2021 - Fish-trace-fossil-KMoisés UgarteNo ratings yet
- Holiday HW - BiologyDocument3 pagesHoliday HW - BiologyAmoghNo ratings yet
- Faunal Exploitation Strategies During The Later Pleistocene in Southern AfricaDocument42 pagesFaunal Exploitation Strategies During The Later Pleistocene in Southern AfricaJerome ReynardNo ratings yet
- Asar - Ecoevorxiv - Preprint ArticuloDocument19 pagesAsar - Ecoevorxiv - Preprint ArticuloNATALIA BEDOYA CORREANo ratings yet
- Diagnostic Aids in Orthodontics FullDocument52 pagesDiagnostic Aids in Orthodontics Fullsunk8650% (2)
- Anatomy of The Inner EarDocument47 pagesAnatomy of The Inner EarAbhilash AntonyNo ratings yet
- c10 Microbiology Tortora TestbankDocument20 pagesc10 Microbiology Tortora Testbankwhitewave25No ratings yet
- Christina and Tim Make A Fart Video by Sellcon2762-D4inko5Document219 pagesChristina and Tim Make A Fart Video by Sellcon2762-D4inko5Jehan DaruwalaNo ratings yet
- Cranial NerveDocument3 pagesCranial NerveBilal SaidNo ratings yet
- 21 Tips To Keep Your Shit Together When You're DepressedDocument3 pages21 Tips To Keep Your Shit Together When You're DepressedPaul MayNo ratings yet
- The Human Body: An Orientation: © 2018 Pearson Education, IncDocument44 pagesThe Human Body: An Orientation: © 2018 Pearson Education, IncShana AquinoNo ratings yet
- Intranet Tdmu Edu Ua Data Kafedra Internal Anatomy Classes SDocument97 pagesIntranet Tdmu Edu Ua Data Kafedra Internal Anatomy Classes SManisanthosh KumarNo ratings yet
- A Soft-Tissue Cephalometric Analysis and Its Use in Orthodontic Treatment Planning. Part IDocument28 pagesA Soft-Tissue Cephalometric Analysis and Its Use in Orthodontic Treatment Planning. Part IJorge Ramírez100% (1)
- Heart Muscle: The Heart As A Pump and FunctionDocument4 pagesHeart Muscle: The Heart As A Pump and Functionmcwnotes100% (1)
- Albert Gunther - Reptiles of IndiaDocument550 pagesAlbert Gunther - Reptiles of IndiaMaximiliano VillarrealNo ratings yet
- Assessment and Diagnostic Tests - IntegumentaryDocument49 pagesAssessment and Diagnostic Tests - IntegumentaryAbdulla AshourNo ratings yet
- AbdomenDocument55 pagesAbdomenAngie LawrenceNo ratings yet
- PhenacetinDocument16 pagesPhenacetinJericSalcedoNo ratings yet
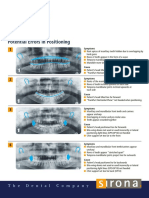
- Potential Errors PositioningDocument2 pagesPotential Errors PositioningROGERQUIROZNo ratings yet
- Module 4 Respiratory SystemDocument8 pagesModule 4 Respiratory SystemJake Donely C. PaduaNo ratings yet
- Consciousness and Hypnagogia PDFDocument30 pagesConsciousness and Hypnagogia PDFprofesorbartolomeo100% (1)
- TUM MIM EssayDocument6 pagesTUM MIM EssayAyeshaAslamNo ratings yet
- Perform Digital VenogramDocument5 pagesPerform Digital VenogramRudy HermosilloNo ratings yet
- Report PDFDocument6 pagesReport PDFAayushiNo ratings yet
- NCP SicuDocument6 pagesNCP SicuChoco MuchoNo ratings yet
- GED-Test-Science (90 Min Questions)Document18 pagesGED-Test-Science (90 Min Questions)Andrew KyawNo ratings yet
- Exalted 3ed Core - Rules - Removed Parts v1Document20 pagesExalted 3ed Core - Rules - Removed Parts v1Rafael SoaresNo ratings yet
- Do All Animals Sleep?: Jerome M. SiegelDocument6 pagesDo All Animals Sleep?: Jerome M. Siegelantoni_gamundi3942No ratings yet
- The Digestive SystemDocument21 pagesThe Digestive SystemjuliusromatolentinoNo ratings yet
- Pathophysiology of Hemolytic AnemiaDocument2 pagesPathophysiology of Hemolytic AnemiaArvin CruzNo ratings yet
- BIOL 105 S 2011 Kidney CH 18 Practice Midterm Exam 2 110429.1Document27 pagesBIOL 105 S 2011 Kidney CH 18 Practice Midterm Exam 2 110429.1NerdyMiggitNo ratings yet
- O.P. Choudhary and PriyankaDocument7 pagesO.P. Choudhary and PriyankaDiary ThalitaNo ratings yet

























































































Download as pdf or txt
You might also like
- Life Orientation Term 3 Grade 11Document11 pagesLife Orientation Term 3 Grade 11farihah75% (4)
- Diabetic Nephropathy Case StudyDocument61 pagesDiabetic Nephropathy Case Studyoxidalaj98% (63)
- EvolutionDocument4 pagesEvolutionChaudhry AzeemNo ratings yet
- From Purgatorius Ceratops To Homo Sapiens: I. Primate Evolutionary HistoryDocument10 pagesFrom Purgatorius Ceratops To Homo Sapiens: I. Primate Evolutionary HistoryquiquebravoNo ratings yet
- SBB 6014 Evolution of MammalsDocument19 pagesSBB 6014 Evolution of MammalsJauha HanafiNo ratings yet
- Article Fossil AnimalsDocument4 pagesArticle Fossil AnimalsmozartNo ratings yet
- Mammal: MammalsDocument13 pagesMammal: Mammalsdaitya99No ratings yet
- Ars Technica - The Paja FormationDocument4 pagesArs Technica - The Paja Formationmilton74No ratings yet
- SynapsidsDocument7 pagesSynapsidsapi-407347220No ratings yet
- The Earliest Possible Hominids: Sahelanthropus TchadensisDocument7 pagesThe Earliest Possible Hominids: Sahelanthropus Tchadensisdod_nurNo ratings yet
- PDF Brusatteetal2010TyrannosaurPaleobiologyDocument24 pagesPDF Brusatteetal2010TyrannosaurPaleobiologyArnab DasNo ratings yet
- Evidence of Evolution On Animal VertebrateDocument9 pagesEvidence of Evolution On Animal Vertebratejohanes manaluNo ratings yet
- Septianisa UlfalahDocument12 pagesSeptianisa UlfalahSeptianisa UlfalahNo ratings yet
- Ucsp (Lesson 1)Document28 pagesUcsp (Lesson 1)united nationNo ratings yet
- Chapter 4Document172 pagesChapter 4Aan AkmalNo ratings yet
- UE Evograms Jaws To EarsDocument3 pagesUE Evograms Jaws To Earsskline3No ratings yet
- Dinosaur: Dinosaurs Are A Diverse Group of Animals of TheDocument5 pagesDinosaur: Dinosaurs Are A Diverse Group of Animals of ThebabublsNo ratings yet
- Dino Jaws Information SheetsDocument55 pagesDino Jaws Information SheetsJacob PruittNo ratings yet
- Evolution Epidemiology and Etiology of Temporomandibular Joint DisordersDocument6 pagesEvolution Epidemiology and Etiology of Temporomandibular Joint DisordersCM Panda CedeesNo ratings yet
- App 20100005Document22 pagesApp 20100005Ghaier KazmiNo ratings yet
- Life-History Evolution in ReptilesDocument11 pagesLife-History Evolution in ReptilesjeraldNo ratings yet
- Anthropological Concepts in Clinical OrthodonticsDocument42 pagesAnthropological Concepts in Clinical Orthodonticsayesha rathore100% (1)
- Freeman. 1981. Correspondence of Food Habits and Morphology in Insectivorous BatsDocument9 pagesFreeman. 1981. Correspondence of Food Habits and Morphology in Insectivorous BatsHenry CondoriNo ratings yet
- 216) Escanear0001Document1 page216) Escanear0001Beatriz MogollonNo ratings yet
- 216) Escanear0001Document1 page216) Escanear0001Beatriz MogollonNo ratings yet
- Bed 130 - Week 1 TaskDocument27 pagesBed 130 - Week 1 TaskPaul Ivan L. PazNo ratings yet
- Origins of CetaceansDocument17 pagesOrigins of CetaceansVengadesh Krishna KumarNo ratings yet
- The Origin and Evolution of DinosaursDocument55 pagesThe Origin and Evolution of DinosaursTiago Savignon Cardoso MachadoNo ratings yet
- Biodiversity and EvolutionDocument99 pagesBiodiversity and EvolutionMaria Jamilla R. Pua100% (1)
- AEB 317 Introductory EntomologyDocument35 pagesAEB 317 Introductory EntomologyOgualu FavourNo ratings yet
- Earth History - The Geologic Time ScaleDocument26 pagesEarth History - The Geologic Time ScaleCarminda Subaba CastilloNo ratings yet
- GEN-BIO-2-History of Life On EarthDocument22 pagesGEN-BIO-2-History of Life On EarthGeminiNo ratings yet
- Cope's Rule and The Adaptive Landscape of Dinosaur Body Size EvolutionDocument36 pagesCope's Rule and The Adaptive Landscape of Dinosaur Body Size Evolutionbraydenstoch777No ratings yet
- Etymology: DimetrodonDocument5 pagesEtymology: DimetrodonAjay SinghNo ratings yet
- Fossil Records PDFDocument10 pagesFossil Records PDFShane CubioNo ratings yet
- Paleogene and NeogeneDocument41 pagesPaleogene and Neogenefredcramer081No ratings yet
- Half A Billion Years of BackbonesDocument1 pageHalf A Billion Years of BackbonesM HanifNo ratings yet
- Homo HobbitDocument4 pagesHomo HobbitAndrew JonesNo ratings yet
- Fossils & EvolutionDocument11 pagesFossils & EvolutionDaniel BerryNo ratings yet
- Growth Patterns of Fossil Vertebrates As Deduced From Bone Microstructure: Case Studies From IndiaDocument12 pagesGrowth Patterns of Fossil Vertebrates As Deduced From Bone Microstructure: Case Studies From IndiaOMSURYACHANDRASIVANo ratings yet
- #2 Chapter IIDocument17 pages#2 Chapter IIAngelyn Bantilo SerronaNo ratings yet
- Sauropod Dinosaur FossilDocument3 pagesSauropod Dinosaur FossilWestern Ang'ong'aNo ratings yet
- Darwin's Theory EssayDocument6 pagesDarwin's Theory EssayLogkeetha SarckunadasanNo ratings yet
- Bernard ReviewerDocument2 pagesBernard ReviewerBernard BrowneNo ratings yet
- The Existence of TrilobitesDocument4 pagesThe Existence of Trilobitesvidur_talrejaNo ratings yet
- The Origin of Animal Body PlansDocument15 pagesThe Origin of Animal Body PlansDiego LoboNo ratings yet
- Diet and The Evolution of The Earliest Human Ancestors: Mark F. Teaford and Peter S. UngarDocument6 pagesDiet and The Evolution of The Earliest Human Ancestors: Mark F. Teaford and Peter S. UngarRaul Diaz TorresNo ratings yet
- Rocio Huizar Instructor Teri Potter Anthropology 1020-012Document5 pagesRocio Huizar Instructor Teri Potter Anthropology 1020-012api-311470705No ratings yet
- Macroevolution and The Formation of New SpeciesDocument19 pagesMacroevolution and The Formation of New Speciesmichelle ann sabilonaNo ratings yet
- Haas 1998Document33 pagesHaas 1998lassaad.el.hanafiNo ratings yet
- UNIT 1palaeontology Handout..Document8 pagesUNIT 1palaeontology Handout..getnet gizachewNo ratings yet
- Lesson 5.1 SummaryDocument4 pagesLesson 5.1 SummaryJaime MallaNo ratings yet
- Overview of ProjectDocument14 pagesOverview of Projectofmephotos4No ratings yet
- Population Structure of The Gomphothere Stegomastodon Waringi (Mammalia: Proboscidea: Gomphotheriidae) From The Pleistocene of BrazilDocument14 pagesPopulation Structure of The Gomphothere Stegomastodon Waringi (Mammalia: Proboscidea: Gomphotheriidae) From The Pleistocene of BrazilKanekoNo ratings yet
- Blueprint of LifeDocument11 pagesBlueprint of Lifetimeflies23No ratings yet
- General Description: Triceratops Megalosaurus IguanodonDocument31 pagesGeneral Description: Triceratops Megalosaurus IguanodonRicardo Daniel Aranda ApolinarioNo ratings yet
- 4-Rodriguez-De La Rosa - Etal - 2021 - Fish-trace-fossil-KDocument10 pages4-Rodriguez-De La Rosa - Etal - 2021 - Fish-trace-fossil-KMoisés UgarteNo ratings yet
- Holiday HW - BiologyDocument3 pagesHoliday HW - BiologyAmoghNo ratings yet
- Faunal Exploitation Strategies During The Later Pleistocene in Southern AfricaDocument42 pagesFaunal Exploitation Strategies During The Later Pleistocene in Southern AfricaJerome ReynardNo ratings yet
- Asar - Ecoevorxiv - Preprint ArticuloDocument19 pagesAsar - Ecoevorxiv - Preprint ArticuloNATALIA BEDOYA CORREANo ratings yet
- Diagnostic Aids in Orthodontics FullDocument52 pagesDiagnostic Aids in Orthodontics Fullsunk8650% (2)
- Anatomy of The Inner EarDocument47 pagesAnatomy of The Inner EarAbhilash AntonyNo ratings yet
- c10 Microbiology Tortora TestbankDocument20 pagesc10 Microbiology Tortora Testbankwhitewave25No ratings yet
- Christina and Tim Make A Fart Video by Sellcon2762-D4inko5Document219 pagesChristina and Tim Make A Fart Video by Sellcon2762-D4inko5Jehan DaruwalaNo ratings yet
- Cranial NerveDocument3 pagesCranial NerveBilal SaidNo ratings yet
- 21 Tips To Keep Your Shit Together When You're DepressedDocument3 pages21 Tips To Keep Your Shit Together When You're DepressedPaul MayNo ratings yet
- The Human Body: An Orientation: © 2018 Pearson Education, IncDocument44 pagesThe Human Body: An Orientation: © 2018 Pearson Education, IncShana AquinoNo ratings yet
- Intranet Tdmu Edu Ua Data Kafedra Internal Anatomy Classes SDocument97 pagesIntranet Tdmu Edu Ua Data Kafedra Internal Anatomy Classes SManisanthosh KumarNo ratings yet
- A Soft-Tissue Cephalometric Analysis and Its Use in Orthodontic Treatment Planning. Part IDocument28 pagesA Soft-Tissue Cephalometric Analysis and Its Use in Orthodontic Treatment Planning. Part IJorge Ramírez100% (1)
- Heart Muscle: The Heart As A Pump and FunctionDocument4 pagesHeart Muscle: The Heart As A Pump and Functionmcwnotes100% (1)
- Albert Gunther - Reptiles of IndiaDocument550 pagesAlbert Gunther - Reptiles of IndiaMaximiliano VillarrealNo ratings yet
- Assessment and Diagnostic Tests - IntegumentaryDocument49 pagesAssessment and Diagnostic Tests - IntegumentaryAbdulla AshourNo ratings yet
- AbdomenDocument55 pagesAbdomenAngie LawrenceNo ratings yet
- PhenacetinDocument16 pagesPhenacetinJericSalcedoNo ratings yet
- Potential Errors PositioningDocument2 pagesPotential Errors PositioningROGERQUIROZNo ratings yet
- Module 4 Respiratory SystemDocument8 pagesModule 4 Respiratory SystemJake Donely C. PaduaNo ratings yet
- Consciousness and Hypnagogia PDFDocument30 pagesConsciousness and Hypnagogia PDFprofesorbartolomeo100% (1)
- TUM MIM EssayDocument6 pagesTUM MIM EssayAyeshaAslamNo ratings yet
- Perform Digital VenogramDocument5 pagesPerform Digital VenogramRudy HermosilloNo ratings yet
- Report PDFDocument6 pagesReport PDFAayushiNo ratings yet
- NCP SicuDocument6 pagesNCP SicuChoco MuchoNo ratings yet
- GED-Test-Science (90 Min Questions)Document18 pagesGED-Test-Science (90 Min Questions)Andrew KyawNo ratings yet
- Exalted 3ed Core - Rules - Removed Parts v1Document20 pagesExalted 3ed Core - Rules - Removed Parts v1Rafael SoaresNo ratings yet
- Do All Animals Sleep?: Jerome M. SiegelDocument6 pagesDo All Animals Sleep?: Jerome M. Siegelantoni_gamundi3942No ratings yet
- The Digestive SystemDocument21 pagesThe Digestive SystemjuliusromatolentinoNo ratings yet
- Pathophysiology of Hemolytic AnemiaDocument2 pagesPathophysiology of Hemolytic AnemiaArvin CruzNo ratings yet
- BIOL 105 S 2011 Kidney CH 18 Practice Midterm Exam 2 110429.1Document27 pagesBIOL 105 S 2011 Kidney CH 18 Practice Midterm Exam 2 110429.1NerdyMiggitNo ratings yet
- O.P. Choudhary and PriyankaDocument7 pagesO.P. Choudhary and PriyankaDiary ThalitaNo ratings yet