Download as pdf or txt
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5822)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1093)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (852)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (898)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (349)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (403)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Mec32 HW3Document4 pagesMec32 HW3Isabelle LunaNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- NDT 30P Penetrant Testing 090217 - Notes For Electronic DistributionDocument105 pagesNDT 30P Penetrant Testing 090217 - Notes For Electronic DistributionUlvi Ismayilov100% (6)
- Pre Insulated Pipe SystemDocument8 pagesPre Insulated Pipe SystemCraciun DanielNo ratings yet
- Conferinte Sectiunea I - cnc2018Document15 pagesConferinte Sectiunea I - cnc2018Cristina IoanaNo ratings yet
- Casting and Welding TrainingDocument120 pagesCasting and Welding TrainingMohd SamNo ratings yet
- Nichrome60 Wire Data SheetDocument2 pagesNichrome60 Wire Data SheetvvingtsabtaNo ratings yet
- Take Test: SW 3BDocument4 pagesTake Test: SW 3BJoseph ZamoraNo ratings yet
- 1516-Advanced Paper-2 Set-A PDFDocument21 pages1516-Advanced Paper-2 Set-A PDFGame CrashNo ratings yet
- Bibliografie CaeDocument1 pageBibliografie CaeBogdanel BucurNo ratings yet
- Mud ViscosityDocument5 pagesMud Viscosityhindn162No ratings yet
- Iit Jee Chemistry DPP by :pjoyDocument3 pagesIit Jee Chemistry DPP by :pjoyPrakash Joy50% (4)
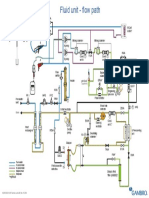
- Ak 96 Fluid Unit - Flow Path: (Upgrade Kit)Document1 pageAk 96 Fluid Unit - Flow Path: (Upgrade Kit)Mounir AngadNo ratings yet
- Msds PDFDocument5 pagesMsds PDFputri azizahNo ratings yet
- ISO Grid SalmonellaDocument1 pageISO Grid Salmonellaapi-3697331No ratings yet
- Development of A Validated RP HPLC DAD MDocument8 pagesDevelopment of A Validated RP HPLC DAD Medlyntonic2No ratings yet
- ORMES - Orbitally Rearranged Monatomic Elements: Superconductivity and Modern AlchemyDocument38 pagesORMES - Orbitally Rearranged Monatomic Elements: Superconductivity and Modern AlchemyBytalesNo ratings yet
- Cement IntroDocument29 pagesCement IntroNitish KumarNo ratings yet
- CP 10 - Factors Affectinging PhotosynthesisDocument3 pagesCP 10 - Factors Affectinging Photosynthesisrifu91No ratings yet
- GHS Lauramine Oxide MSDSDocument7 pagesGHS Lauramine Oxide MSDSAnup KumarNo ratings yet
- CBLM FloresDocument42 pagesCBLM FloresJinky Aydalla100% (2)
- Application Procedure For Interzone 505 and Interthane 990 SystemDocument16 pagesApplication Procedure For Interzone 505 and Interthane 990 SystemMehman NasibovNo ratings yet
- Assignment Cat 1Document2 pagesAssignment Cat 1ppraveenksrceNo ratings yet
- Narayana 06-06-2022 - Outgoing SR - Jee Main Model Gtm-6 - SolDocument16 pagesNarayana 06-06-2022 - Outgoing SR - Jee Main Model Gtm-6 - SolYuva AkhilNo ratings yet
- Passos Et Al. - 2017 - Towards Energy Neutral Microalgae-Based Wastewater Treatment PlantsDocument9 pagesPassos Et Al. - 2017 - Towards Energy Neutral Microalgae-Based Wastewater Treatment PlantsfvassisNo ratings yet
- 69 Polygalacturonases: Jacques A. E. Benen Jaap VisserDocument10 pages69 Polygalacturonases: Jacques A. E. Benen Jaap VisserElena Raluca BanescuNo ratings yet
- AIATS Schedule For RM (XII Passed) 2023-24Document2 pagesAIATS Schedule For RM (XII Passed) 2023-24sampadaNo ratings yet
- Fundamentals of Multiphase Flow: Prof. G. DasDocument34 pagesFundamentals of Multiphase Flow: Prof. G. DasVRUSHABHNo ratings yet
- Electron Impact Ionization (EI)Document3 pagesElectron Impact Ionization (EI)محمد ازواديNo ratings yet
- Kelompok 8 - Irsan Bagas Maulana HarahapDocument20 pagesKelompok 8 - Irsan Bagas Maulana HarahapGale HawthorneNo ratings yet
- (Oklahoma Notes) Roger Thies Ph.D. (Auth.), Roger Thies Ph.D. (Eds.) - Physiology-Springer-Verlag New York (1995)Document287 pages(Oklahoma Notes) Roger Thies Ph.D. (Auth.), Roger Thies Ph.D. (Eds.) - Physiology-Springer-Verlag New York (1995)MaadaNo ratings yet