
Assignment On Regulation of Transcription, Lytic Lysogeny Cascade and SOS Regulatory System
Assignment On Regulation of Transcription, Lytic Lysogeny Cascade and SOS Regulatory System
You might also like
- SCIN130 Week 5 QuizDocument22 pagesSCIN130 Week 5 Quiznew rhondaldNo ratings yet
- Pro TranscriptionDocument20 pagesPro TranscriptionnitralekhaNo ratings yet
- Promoters and Plasmids PDFDocument4 pagesPromoters and Plasmids PDFAndika MardiantoNo ratings yet
- Enhancer: Q1) What Is Role of Enhancers and Promoters in Transcription of Eukaryotes? AnsDocument9 pagesEnhancer: Q1) What Is Role of Enhancers and Promoters in Transcription of Eukaryotes? AnsSudeep BiswasNo ratings yet
- 11TrpAMO LicyayoDocument8 pages11TrpAMO LicyayoMohamidin MamalapatNo ratings yet
- TranscriptionDocument21 pagesTranscriptionYehya JaberNo ratings yet
- Molecular Biology NotesDocument8 pagesMolecular Biology Notesbyunus88No ratings yet
- Transcriprion ManuscriptDocument14 pagesTranscriprion ManuscriptAsif AhmedNo ratings yet
- Science FairDocument41 pagesScience FairSafanaNo ratings yet
- TranscriptionDocument10 pagesTranscriptionHardik ManekNo ratings yet
- SYBT TranscriptionDocument69 pagesSYBT TranscriptionMeir SabooNo ratings yet
- Messenger Rnas (Mrnas) : This Class of Rnas Are The Genetic Coding TemplatesDocument8 pagesMessenger Rnas (Mrnas) : This Class of Rnas Are The Genetic Coding TemplatesRavi AlugubelliNo ratings yet
- Lec 7 TranscriptionDocument30 pagesLec 7 TranscriptionYasmin BalochNo ratings yet
- RNA Synthesis, Processing SBT 211Document59 pagesRNA Synthesis, Processing SBT 211TADIWANASHE TINONETSANANo ratings yet
- Gene ExpressionDocument19 pagesGene Expressionkvicto100% (1)
- E.coli RNA PolymeraseDocument17 pagesE.coli RNA PolymeraseM.PRASAD NAIDUNo ratings yet
- DNA TranscriptionDocument25 pagesDNA TranscriptionAkss ShauryaNo ratings yet
- Transcription in EukaryotesDocument49 pagesTranscription in EukaryotesmrlabdesigneNo ratings yet
- Transcription and Regulation of Gene Expression: By: Lyka Marie C. Falcasantos BSN - 1DDocument16 pagesTranscription and Regulation of Gene Expression: By: Lyka Marie C. Falcasantos BSN - 1DRemzAbdullaNo ratings yet
- Lecture 2 - Transcription and RNA ProcessingDocument22 pagesLecture 2 - Transcription and RNA ProcessingErika KulićNo ratings yet
- Transcription Prokaryotes 2012-cDocument27 pagesTranscription Prokaryotes 2012-cAnupama PatiNo ratings yet
- Dna L12 NotesDocument6 pagesDna L12 NotesellieNo ratings yet
- RNA Structure, Function, and Synthesis RNADocument6 pagesRNA Structure, Function, and Synthesis RNAMohini BajajNo ratings yet
- DNA Transcription - Learn Science at ScitableDocument4 pagesDNA Transcription - Learn Science at ScitableSheryl Lin Dayrit DolorNo ratings yet
- Transcription in Prokaryotes: Single RNA PolymeraseDocument29 pagesTranscription in Prokaryotes: Single RNA PolymeraseShubhamNo ratings yet
- Molecular Biology Lecture 11Document38 pagesMolecular Biology Lecture 11Anton IrvingNo ratings yet
- Decker 2Document7 pagesDecker 2Katarina Kaca GacevicNo ratings yet
- Gene Expression and DNA ReplicationDocument22 pagesGene Expression and DNA ReplicationHagar AlaaNo ratings yet
- RNA ReplicationDocument23 pagesRNA ReplicationDharaneeshwari Siva-F&NNo ratings yet
- Surgery of Transcription InsightDocument19 pagesSurgery of Transcription Insightnhung_ducuments8261No ratings yet
- Molecular Microbiology MAP Prep FinalDocument41 pagesMolecular Microbiology MAP Prep FinalAdi SassonNo ratings yet
- 25 LN RNA Organization and Transcription CNRAADocument55 pages25 LN RNA Organization and Transcription CNRAADakshitha DharmakeerthiNo ratings yet
- Transcription ProkaryoticDocument30 pagesTranscription ProkaryoticDibya Jyoti ParidaNo ratings yet
- Transcription and RNA Processing in EukaryotesDocument13 pagesTranscription and RNA Processing in EukaryotesAbdulfattah NoorNo ratings yet
- Prokaryotes and EukaryoticDocument8 pagesProkaryotes and EukaryoticYasirNo ratings yet
- Transcription WorkingDocument51 pagesTranscription Workingapi-3858544No ratings yet
- TranscriptionDocument56 pagesTranscriptionVipin100% (8)
- RNA Synthesis, Processing & ModificationDocument53 pagesRNA Synthesis, Processing & Modificationamanialwerfalli4No ratings yet
- Report in Cell BiologyIIDocument3 pagesReport in Cell BiologyIIGenessa Agustin BuenafeNo ratings yet
- Molecular Biology (Genes and Mutations)Document61 pagesMolecular Biology (Genes and Mutations)Justin PRUDENCIONo ratings yet
- Transcription (Semister I MIC)Document117 pagesTranscription (Semister I MIC)Challagandla AnilNo ratings yet
- TranscriptionDocument34 pagesTranscriptiondrhydrogenNo ratings yet
- Transcription in ProkaryotesDocument18 pagesTranscription in ProkaryotesPrakash100% (1)
- Transcription EukarDocument3 pagesTranscription EukarnitralekhaNo ratings yet
- TranscriptionDocument61 pagesTranscriptiondeepak mauryaNo ratings yet
- Transcription: Department of Biochemistry Susan B. Caceres-Tengco, M.DDocument40 pagesTranscription: Department of Biochemistry Susan B. Caceres-Tengco, M.DLen ArellanoNo ratings yet
- Transcription Part 1Document5 pagesTranscription Part 1John NyimbiliNo ratings yet
- 4 Gene ExpressionDocument35 pages4 Gene ExpressionThảo ThảoNo ratings yet
- Gene Regulation (Post Translation)Document111 pagesGene Regulation (Post Translation)raryanraj44No ratings yet
- Meredith Barb Homework-2 (25 Points) Due On Sept. 24 by 5:00pmDocument5 pagesMeredith Barb Homework-2 (25 Points) Due On Sept. 24 by 5:00pmMeredith BarbNo ratings yet
- Transkripsi Dan Translasi SMA Permata Insani IslamicDocument58 pagesTranskripsi Dan Translasi SMA Permata Insani IslamicMaria Angelina Genere KobanNo ratings yet
- L1 Expression vectorsDocument14 pagesL1 Expression vectorsMaxwel OdongoNo ratings yet
- Transcripción - 1 Bio 363 Genética Molecular I Semestre 2021 Sergio MarshallDocument43 pagesTranscripción - 1 Bio 363 Genética Molecular I Semestre 2021 Sergio MarshallIgnacioHandyMoralesNo ratings yet
- Chemistry Lec.22 (Nucleic Acids 6)Document6 pagesChemistry Lec.22 (Nucleic Acids 6)Muhammed AbdulsamadNo ratings yet
- Transcription: - by - S. Sivaranjani Arunnehru - Assistant Professor - Bon Secours College For Women - ThanjavurDocument33 pagesTranscription: - by - S. Sivaranjani Arunnehru - Assistant Professor - Bon Secours College For Women - ThanjavurGayathri deviNo ratings yet
- Rna and Amino AcidDocument14 pagesRna and Amino AcidIndestructible queenNo ratings yet
- Mrs. Ofelia Solano Saludar: Department of Natural Sciences University of St. La Salle Bacolod CityDocument55 pagesMrs. Ofelia Solano Saludar: Department of Natural Sciences University of St. La Salle Bacolod CitymskikiNo ratings yet
- How The Genes Are Regulated & Expressed?: AssignmentDocument17 pagesHow The Genes Are Regulated & Expressed?: AssignmentSuraiya Siraj NehaNo ratings yet
- Regulation of Gene Expression in Eukaryotes: Presented by Quratulain (19-Arid-1403) Asadullah (19-Arid-1379)Document32 pagesRegulation of Gene Expression in Eukaryotes: Presented by Quratulain (19-Arid-1403) Asadullah (19-Arid-1379)Beauty LiciousNo ratings yet
- Molecular Biology: TranscriptionDocument101 pagesMolecular Biology: TranscriptionMarni Arta Uli SimbolonNo ratings yet
- Must Review ThisDocument19 pagesMust Review Thispmp123456No ratings yet
- DLP - Tomatom. Science 10. q3 - DNA REPLICAtionDocument10 pagesDLP - Tomatom. Science 10. q3 - DNA REPLICAtionalchristian tomatomNo ratings yet
- Cytogenetics Final ExamDocument20 pagesCytogenetics Final ExamMICHAEL JOHN AGUILARNo ratings yet
- Genetics and Cell Biology Exam 1 Study GuideDocument7 pagesGenetics and Cell Biology Exam 1 Study GuideSamuel JosiahNo ratings yet
- DNA Replication Study GuideDocument3 pagesDNA Replication Study Guide黄诗雨No ratings yet
- Ds83-Replication Review Formative AssessmentDocument4 pagesDs83-Replication Review Formative Assessmentapi-110789702No ratings yet
- Lecture 6. Genetic 2Document85 pagesLecture 6. Genetic 2Thanhh ThaooNo ratings yet
- Eukaryotic Gene Transcription NotesDocument19 pagesEukaryotic Gene Transcription NotesEshaNo ratings yet
- Science Assessment TaskDocument4 pagesScience Assessment TaskNabiha SyedaNo ratings yet
- Chapter 25 DNA Metabolism: Replication (Faithful Reproduction) - Which Must Be IncrediblyDocument28 pagesChapter 25 DNA Metabolism: Replication (Faithful Reproduction) - Which Must Be IncrediblyRainidah Mangotara Ismael-DericoNo ratings yet
- Eukaryotic ReplisomeDocument344 pagesEukaryotic ReplisomeIsmaelVázquezEspinoza100% (1)
- Test Bank For Textbook of Biochemistry With Clinical Correlations 7th Edition Thomas M Devlin DownloadDocument23 pagesTest Bank For Textbook of Biochemistry With Clinical Correlations 7th Edition Thomas M Devlin DownloadMeganGomezpizb100% (20)
- Eukaryotic DNA Polymerases: Sue Cotterill, Stephen KearseyDocument6 pagesEukaryotic DNA Polymerases: Sue Cotterill, Stephen KearseyOphy FirmansyahNo ratings yet
- 13-1: The Genetic Material: Frederick Griffith's Experiment - TransformationDocument7 pages13-1: The Genetic Material: Frederick Griffith's Experiment - Transformationapi-233187566No ratings yet
- BIO 314 - Final ExamDocument27 pagesBIO 314 - Final ExamNerdy Notes Inc.100% (1)
- DNA Replication Escape Room DocumentsDocument42 pagesDNA Replication Escape Room DocumentsVictoria WarrenNo ratings yet
- 3 Bacterial Genetics I & II DentistryDocument90 pages3 Bacterial Genetics I & II DentistryMajd HallakNo ratings yet
- Molecular Medicine QuestionsDocument2 pagesMolecular Medicine QuestionsJordan ConstantineNo ratings yet
- Central Dogma of Molecular Biology RendraDocument36 pagesCentral Dogma of Molecular Biology RendraYuliaji Narendra PutraNo ratings yet
- Kinetoplast DNA Replication: F. Torri, Laura Rocco Carpenter, and Paul T. EnglundDocument14 pagesKinetoplast DNA Replication: F. Torri, Laura Rocco Carpenter, and Paul T. EnglundAdrian OrtegaNo ratings yet
- DNA ReplicationDocument13 pagesDNA ReplicationJoanne NgauNo ratings yet
- DNA Replication NotesDocument2 pagesDNA Replication NotesMart Tin100% (1)
- Lecture NotesDocument32 pagesLecture Noteslira shresthaNo ratings yet
- Takano 1994Document6 pagesTakano 1994maryjoseNo ratings yet
- Biochemistry MCQs 2Document18 pagesBiochemistry MCQs 2MUHAMMAD NAEEM IQBALNo ratings yet
- Mmoi BC SbaDocument15 pagesMmoi BC SbaYoon YoonNo ratings yet
- Chapter 16 - Transcription, RNA Processing, and TranslationDocument18 pagesChapter 16 - Transcription, RNA Processing, and TranslationRaviNagiNo ratings yet
- Cytogenetics Seatwork No. 4 DNA ReplicationDocument4 pagesCytogenetics Seatwork No. 4 DNA ReplicationKemverly RanaNo ratings yet
- DNA Repair From Lehninger - Addtnlnotes PDFDocument13 pagesDNA Repair From Lehninger - Addtnlnotes PDFAlthea Karmylle M. BonitaNo ratings yet



















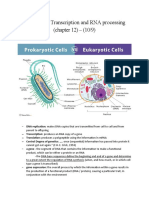





































































Download as doc, pdf, or txt
You might also like
- SCIN130 Week 5 QuizDocument22 pagesSCIN130 Week 5 Quiznew rhondaldNo ratings yet
- Pro TranscriptionDocument20 pagesPro TranscriptionnitralekhaNo ratings yet
- Promoters and Plasmids PDFDocument4 pagesPromoters and Plasmids PDFAndika MardiantoNo ratings yet
- Enhancer: Q1) What Is Role of Enhancers and Promoters in Transcription of Eukaryotes? AnsDocument9 pagesEnhancer: Q1) What Is Role of Enhancers and Promoters in Transcription of Eukaryotes? AnsSudeep BiswasNo ratings yet
- 11TrpAMO LicyayoDocument8 pages11TrpAMO LicyayoMohamidin MamalapatNo ratings yet
- TranscriptionDocument21 pagesTranscriptionYehya JaberNo ratings yet
- Molecular Biology NotesDocument8 pagesMolecular Biology Notesbyunus88No ratings yet
- Transcriprion ManuscriptDocument14 pagesTranscriprion ManuscriptAsif AhmedNo ratings yet
- Science FairDocument41 pagesScience FairSafanaNo ratings yet
- TranscriptionDocument10 pagesTranscriptionHardik ManekNo ratings yet
- SYBT TranscriptionDocument69 pagesSYBT TranscriptionMeir SabooNo ratings yet
- Messenger Rnas (Mrnas) : This Class of Rnas Are The Genetic Coding TemplatesDocument8 pagesMessenger Rnas (Mrnas) : This Class of Rnas Are The Genetic Coding TemplatesRavi AlugubelliNo ratings yet
- Lec 7 TranscriptionDocument30 pagesLec 7 TranscriptionYasmin BalochNo ratings yet
- RNA Synthesis, Processing SBT 211Document59 pagesRNA Synthesis, Processing SBT 211TADIWANASHE TINONETSANANo ratings yet
- Gene ExpressionDocument19 pagesGene Expressionkvicto100% (1)
- E.coli RNA PolymeraseDocument17 pagesE.coli RNA PolymeraseM.PRASAD NAIDUNo ratings yet
- DNA TranscriptionDocument25 pagesDNA TranscriptionAkss ShauryaNo ratings yet
- Transcription in EukaryotesDocument49 pagesTranscription in EukaryotesmrlabdesigneNo ratings yet
- Transcription and Regulation of Gene Expression: By: Lyka Marie C. Falcasantos BSN - 1DDocument16 pagesTranscription and Regulation of Gene Expression: By: Lyka Marie C. Falcasantos BSN - 1DRemzAbdullaNo ratings yet
- Lecture 2 - Transcription and RNA ProcessingDocument22 pagesLecture 2 - Transcription and RNA ProcessingErika KulićNo ratings yet
- Transcription Prokaryotes 2012-cDocument27 pagesTranscription Prokaryotes 2012-cAnupama PatiNo ratings yet
- Dna L12 NotesDocument6 pagesDna L12 NotesellieNo ratings yet
- RNA Structure, Function, and Synthesis RNADocument6 pagesRNA Structure, Function, and Synthesis RNAMohini BajajNo ratings yet
- DNA Transcription - Learn Science at ScitableDocument4 pagesDNA Transcription - Learn Science at ScitableSheryl Lin Dayrit DolorNo ratings yet
- Transcription in Prokaryotes: Single RNA PolymeraseDocument29 pagesTranscription in Prokaryotes: Single RNA PolymeraseShubhamNo ratings yet
- Molecular Biology Lecture 11Document38 pagesMolecular Biology Lecture 11Anton IrvingNo ratings yet
- Decker 2Document7 pagesDecker 2Katarina Kaca GacevicNo ratings yet
- Gene Expression and DNA ReplicationDocument22 pagesGene Expression and DNA ReplicationHagar AlaaNo ratings yet
- RNA ReplicationDocument23 pagesRNA ReplicationDharaneeshwari Siva-F&NNo ratings yet
- Surgery of Transcription InsightDocument19 pagesSurgery of Transcription Insightnhung_ducuments8261No ratings yet
- Molecular Microbiology MAP Prep FinalDocument41 pagesMolecular Microbiology MAP Prep FinalAdi SassonNo ratings yet
- 25 LN RNA Organization and Transcription CNRAADocument55 pages25 LN RNA Organization and Transcription CNRAADakshitha DharmakeerthiNo ratings yet
- Transcription ProkaryoticDocument30 pagesTranscription ProkaryoticDibya Jyoti ParidaNo ratings yet
- Transcription and RNA Processing in EukaryotesDocument13 pagesTranscription and RNA Processing in EukaryotesAbdulfattah NoorNo ratings yet
- Prokaryotes and EukaryoticDocument8 pagesProkaryotes and EukaryoticYasirNo ratings yet
- Transcription WorkingDocument51 pagesTranscription Workingapi-3858544No ratings yet
- TranscriptionDocument56 pagesTranscriptionVipin100% (8)
- RNA Synthesis, Processing & ModificationDocument53 pagesRNA Synthesis, Processing & Modificationamanialwerfalli4No ratings yet
- Report in Cell BiologyIIDocument3 pagesReport in Cell BiologyIIGenessa Agustin BuenafeNo ratings yet
- Molecular Biology (Genes and Mutations)Document61 pagesMolecular Biology (Genes and Mutations)Justin PRUDENCIONo ratings yet
- Transcription (Semister I MIC)Document117 pagesTranscription (Semister I MIC)Challagandla AnilNo ratings yet
- TranscriptionDocument34 pagesTranscriptiondrhydrogenNo ratings yet
- Transcription in ProkaryotesDocument18 pagesTranscription in ProkaryotesPrakash100% (1)
- Transcription EukarDocument3 pagesTranscription EukarnitralekhaNo ratings yet
- TranscriptionDocument61 pagesTranscriptiondeepak mauryaNo ratings yet
- Transcription: Department of Biochemistry Susan B. Caceres-Tengco, M.DDocument40 pagesTranscription: Department of Biochemistry Susan B. Caceres-Tengco, M.DLen ArellanoNo ratings yet
- Transcription Part 1Document5 pagesTranscription Part 1John NyimbiliNo ratings yet
- 4 Gene ExpressionDocument35 pages4 Gene ExpressionThảo ThảoNo ratings yet
- Gene Regulation (Post Translation)Document111 pagesGene Regulation (Post Translation)raryanraj44No ratings yet
- Meredith Barb Homework-2 (25 Points) Due On Sept. 24 by 5:00pmDocument5 pagesMeredith Barb Homework-2 (25 Points) Due On Sept. 24 by 5:00pmMeredith BarbNo ratings yet
- Transkripsi Dan Translasi SMA Permata Insani IslamicDocument58 pagesTranskripsi Dan Translasi SMA Permata Insani IslamicMaria Angelina Genere KobanNo ratings yet
- L1 Expression vectorsDocument14 pagesL1 Expression vectorsMaxwel OdongoNo ratings yet
- Transcripción - 1 Bio 363 Genética Molecular I Semestre 2021 Sergio MarshallDocument43 pagesTranscripción - 1 Bio 363 Genética Molecular I Semestre 2021 Sergio MarshallIgnacioHandyMoralesNo ratings yet
- Chemistry Lec.22 (Nucleic Acids 6)Document6 pagesChemistry Lec.22 (Nucleic Acids 6)Muhammed AbdulsamadNo ratings yet
- Transcription: - by - S. Sivaranjani Arunnehru - Assistant Professor - Bon Secours College For Women - ThanjavurDocument33 pagesTranscription: - by - S. Sivaranjani Arunnehru - Assistant Professor - Bon Secours College For Women - ThanjavurGayathri deviNo ratings yet
- Rna and Amino AcidDocument14 pagesRna and Amino AcidIndestructible queenNo ratings yet
- Mrs. Ofelia Solano Saludar: Department of Natural Sciences University of St. La Salle Bacolod CityDocument55 pagesMrs. Ofelia Solano Saludar: Department of Natural Sciences University of St. La Salle Bacolod CitymskikiNo ratings yet
- How The Genes Are Regulated & Expressed?: AssignmentDocument17 pagesHow The Genes Are Regulated & Expressed?: AssignmentSuraiya Siraj NehaNo ratings yet
- Regulation of Gene Expression in Eukaryotes: Presented by Quratulain (19-Arid-1403) Asadullah (19-Arid-1379)Document32 pagesRegulation of Gene Expression in Eukaryotes: Presented by Quratulain (19-Arid-1403) Asadullah (19-Arid-1379)Beauty LiciousNo ratings yet
- Molecular Biology: TranscriptionDocument101 pagesMolecular Biology: TranscriptionMarni Arta Uli SimbolonNo ratings yet
- Must Review ThisDocument19 pagesMust Review Thispmp123456No ratings yet
- DLP - Tomatom. Science 10. q3 - DNA REPLICAtionDocument10 pagesDLP - Tomatom. Science 10. q3 - DNA REPLICAtionalchristian tomatomNo ratings yet
- Cytogenetics Final ExamDocument20 pagesCytogenetics Final ExamMICHAEL JOHN AGUILARNo ratings yet
- Genetics and Cell Biology Exam 1 Study GuideDocument7 pagesGenetics and Cell Biology Exam 1 Study GuideSamuel JosiahNo ratings yet
- DNA Replication Study GuideDocument3 pagesDNA Replication Study Guide黄诗雨No ratings yet
- Ds83-Replication Review Formative AssessmentDocument4 pagesDs83-Replication Review Formative Assessmentapi-110789702No ratings yet
- Lecture 6. Genetic 2Document85 pagesLecture 6. Genetic 2Thanhh ThaooNo ratings yet
- Eukaryotic Gene Transcription NotesDocument19 pagesEukaryotic Gene Transcription NotesEshaNo ratings yet
- Science Assessment TaskDocument4 pagesScience Assessment TaskNabiha SyedaNo ratings yet
- Chapter 25 DNA Metabolism: Replication (Faithful Reproduction) - Which Must Be IncrediblyDocument28 pagesChapter 25 DNA Metabolism: Replication (Faithful Reproduction) - Which Must Be IncrediblyRainidah Mangotara Ismael-DericoNo ratings yet
- Eukaryotic ReplisomeDocument344 pagesEukaryotic ReplisomeIsmaelVázquezEspinoza100% (1)
- Test Bank For Textbook of Biochemistry With Clinical Correlations 7th Edition Thomas M Devlin DownloadDocument23 pagesTest Bank For Textbook of Biochemistry With Clinical Correlations 7th Edition Thomas M Devlin DownloadMeganGomezpizb100% (20)
- Eukaryotic DNA Polymerases: Sue Cotterill, Stephen KearseyDocument6 pagesEukaryotic DNA Polymerases: Sue Cotterill, Stephen KearseyOphy FirmansyahNo ratings yet
- 13-1: The Genetic Material: Frederick Griffith's Experiment - TransformationDocument7 pages13-1: The Genetic Material: Frederick Griffith's Experiment - Transformationapi-233187566No ratings yet
- BIO 314 - Final ExamDocument27 pagesBIO 314 - Final ExamNerdy Notes Inc.100% (1)
- DNA Replication Escape Room DocumentsDocument42 pagesDNA Replication Escape Room DocumentsVictoria WarrenNo ratings yet
- 3 Bacterial Genetics I & II DentistryDocument90 pages3 Bacterial Genetics I & II DentistryMajd HallakNo ratings yet
- Molecular Medicine QuestionsDocument2 pagesMolecular Medicine QuestionsJordan ConstantineNo ratings yet
- Central Dogma of Molecular Biology RendraDocument36 pagesCentral Dogma of Molecular Biology RendraYuliaji Narendra PutraNo ratings yet
- Kinetoplast DNA Replication: F. Torri, Laura Rocco Carpenter, and Paul T. EnglundDocument14 pagesKinetoplast DNA Replication: F. Torri, Laura Rocco Carpenter, and Paul T. EnglundAdrian OrtegaNo ratings yet
- DNA ReplicationDocument13 pagesDNA ReplicationJoanne NgauNo ratings yet
- DNA Replication NotesDocument2 pagesDNA Replication NotesMart Tin100% (1)
- Lecture NotesDocument32 pagesLecture Noteslira shresthaNo ratings yet
- Takano 1994Document6 pagesTakano 1994maryjoseNo ratings yet
- Biochemistry MCQs 2Document18 pagesBiochemistry MCQs 2MUHAMMAD NAEEM IQBALNo ratings yet
- Mmoi BC SbaDocument15 pagesMmoi BC SbaYoon YoonNo ratings yet
- Chapter 16 - Transcription, RNA Processing, and TranslationDocument18 pagesChapter 16 - Transcription, RNA Processing, and TranslationRaviNagiNo ratings yet
- Cytogenetics Seatwork No. 4 DNA ReplicationDocument4 pagesCytogenetics Seatwork No. 4 DNA ReplicationKemverly RanaNo ratings yet
- DNA Repair From Lehninger - Addtnlnotes PDFDocument13 pagesDNA Repair From Lehninger - Addtnlnotes PDFAlthea Karmylle M. BonitaNo ratings yet