
Base Alterations and Base Damage
Base Alterations and Base Damage
You might also like
- Human Molecular Genetics, 4th Edition PDFDocument704 pagesHuman Molecular Genetics, 4th Edition PDFLi Luo100% (5)
- Pro TranscriptionDocument20 pagesPro TranscriptionnitralekhaNo ratings yet
- Methanol Production and UseDocument403 pagesMethanol Production and UseEvin Joy92% (13)
- The Central Dogma of Molecular BiologyDocument4 pagesThe Central Dogma of Molecular BiologyAlvin Fruelda FaaNo ratings yet
- Transcription and RegulationDocument40 pagesTranscription and RegulationRUDRANSH PUJARINo ratings yet
- TranscriptionDocument23 pagesTranscriptionareen fakhouryNo ratings yet
- TranscriptionDocument10 pagesTranscriptionHardik ManekNo ratings yet
- 25 LN RNA Organization and Transcription CNRAADocument55 pages25 LN RNA Organization and Transcription CNRAADakshitha DharmakeerthiNo ratings yet
- G. Gene Expression and Protein SynthesisDocument50 pagesG. Gene Expression and Protein SynthesisMary Rose Bobis VicenteNo ratings yet
- Lecture On Transcription and TranslationDocument47 pagesLecture On Transcription and TranslationAnna Beatrice BautistaNo ratings yet
- DNA Transcription (Part-1) : DR - Ahmed Salim Mohammed PH.D Molecular MicrobiologyDocument49 pagesDNA Transcription (Part-1) : DR - Ahmed Salim Mohammed PH.D Molecular MicrobiologyDrAhmedSalimNo ratings yet
- Webb - DNA 1Document33 pagesWebb - DNA 1monday125No ratings yet
- Gene Expression: Vikrant Joshi F.Y. Biotechnology 6519 Khalsa College AmritsarDocument31 pagesGene Expression: Vikrant Joshi F.Y. Biotechnology 6519 Khalsa College Amritsarvikrant4018No ratings yet
- Topic 10 Transcription Lecture NotesDocument33 pagesTopic 10 Transcription Lecture NoteskambulukatambilaiNo ratings yet
- Nucleic Acid Metabolism TAUDocument38 pagesNucleic Acid Metabolism TAUFaatimah EssaNo ratings yet
- Regulation of Gene Expression in Eukaryotes: Presented by Quratulain (19-Arid-1403) Asadullah (19-Arid-1379)Document32 pagesRegulation of Gene Expression in Eukaryotes: Presented by Quratulain (19-Arid-1403) Asadullah (19-Arid-1379)Beauty LiciousNo ratings yet
- Kuliah 8. Transcription (Oke)Document43 pagesKuliah 8. Transcription (Oke)Sinta DosiNo ratings yet
- 2 TranscriptionDocument51 pages2 Transcriptionmaksumrabbi rumc1999No ratings yet
- Lecture 2Document33 pagesLecture 2SUNDAS FATIMANo ratings yet
- Molecular Biology: TranscriptionDocument101 pagesMolecular Biology: TranscriptionchintyaNo ratings yet
- Central Dogma of Molecular BiologyDocument6 pagesCentral Dogma of Molecular BiologyNiar Ar-RahmahNo ratings yet
- Transcription and TranslationDocument46 pagesTranscription and TranslationEsther SparksNo ratings yet
- Transcription in ProkaryotesDocument16 pagesTranscription in ProkaryotesAditya Kanwal100% (2)
- Transcription and Post-Transcriptional Processing Translation and Post-Translational Protein ProcessingDocument79 pagesTranscription and Post-Transcriptional Processing Translation and Post-Translational Protein ProcessingBcher ChkirNo ratings yet
- RNA Synthesis and ProcessingDocument17 pagesRNA Synthesis and ProcessingInnocent Clifford MaranduNo ratings yet
- Protein SynthesisDocument60 pagesProtein Synthesislucky mbaselaNo ratings yet
- Molecular Biology: TranscriptionDocument101 pagesMolecular Biology: TranscriptiontantyNo ratings yet
- TranscriptionDocument61 pagesTranscriptiondeepak mauryaNo ratings yet
- TranscriptionDocument56 pagesTranscriptionVipin100% (8)
- Transcription: DR Vaishali Dhat Professor, Department of Biochemistry DR DypmcDocument25 pagesTranscription: DR Vaishali Dhat Professor, Department of Biochemistry DR Dypmcnishkarsh chauhanNo ratings yet
- Lecture 11 TranscriptionDocument21 pagesLecture 11 TranscriptionAditi SharmaNo ratings yet
- Transciption and TranslationDocument31 pagesTransciption and TranslationDuduetsang MosalakataneNo ratings yet
- 6 Lecture-TranslationDocument25 pages6 Lecture-Translationmoazzanrana678No ratings yet
- Transcription: From DNA To RNADocument74 pagesTranscription: From DNA To RNAmd habibur rahmanNo ratings yet
- UntitledDocument45 pagesUntitledjemNo ratings yet
- TRANSKRIPSIDocument19 pagesTRANSKRIPSIZakisyarifudintaqiyNo ratings yet
- Sintesis DNADocument32 pagesSintesis DNAZhafran DarwisNo ratings yet
- Genetik MikrobDocument33 pagesGenetik MikrobekasukmawatyNo ratings yet
- Rna Biosynthesis (Transicription)Document33 pagesRna Biosynthesis (Transicription)Alaa AlmajedNo ratings yet
- Inhibition of DNA Replication and TranscriptionDocument111 pagesInhibition of DNA Replication and TranscriptionPranav Kumar PrabhakarNo ratings yet
- Gene ExpressionDocument64 pagesGene ExpressiondesyNo ratings yet
- Gene ExpressionDocument72 pagesGene ExpressionMika ForwardNo ratings yet
- Science FairDocument41 pagesScience FairSafanaNo ratings yet
- Lesson 9Document66 pagesLesson 9Irica Mae CiervoNo ratings yet
- 106 Ceng w10Document62 pages106 Ceng w10mertNo ratings yet
- Chapter 5 Protein SynthesisDocument26 pagesChapter 5 Protein SynthesisT MokshithaNo ratings yet
- 22 (W13) TranscriptionDocument38 pages22 (W13) TranscriptionMUHAMMAD FAHMI MOHD ZAMZANINo ratings yet
- Transcription 2Document66 pagesTranscription 2Safe BoxNo ratings yet
- Ekspresi GenDocument63 pagesEkspresi GenIndraPramanaIdaBagusNo ratings yet
- PROCESS of TRANSKRIPSI, TRANSLASI, REPLIKASI DNA2Document27 pagesPROCESS of TRANSKRIPSI, TRANSLASI, REPLIKASI DNA2istatinNo ratings yet
- TUTORIAL: DNA BIOLOGY and TECHNOLOGY 1. Describe The Biochemical CompositionDocument6 pagesTUTORIAL: DNA BIOLOGY and TECHNOLOGY 1. Describe The Biochemical Compositionaesha89No ratings yet
- Biochemistry: RNA Synthesis and ProcessingDocument54 pagesBiochemistry: RNA Synthesis and ProcessingAqsa YaminNo ratings yet
- Transcription ProkaryoticDocument30 pagesTranscription ProkaryoticDibya Jyoti ParidaNo ratings yet
- RNA Processing or Post Transcriptional Modifications.Document23 pagesRNA Processing or Post Transcriptional Modifications.saeed313bbtNo ratings yet
- DNA-Dependent Synthesis of RNA: (Transcription)Document10 pagesDNA-Dependent Synthesis of RNA: (Transcription)Shivam SharmaNo ratings yet
- TranscriptionDocument34 pagesTranscriptiondrhydrogenNo ratings yet
- Lecture 5 Diagnostic GenomicsDocument33 pagesLecture 5 Diagnostic Genomicssales zfNo ratings yet
- Lect 4 Prokaryotic Gene StructureDocument45 pagesLect 4 Prokaryotic Gene StructureElias LibayNo ratings yet
- TransDocument29 pagesTransDrLakshmi Priya ThyagarajanNo ratings yet
- Chapter 6 SummaryDocument28 pagesChapter 6 SummaryCharlotteNo ratings yet
- Telomerases: Chemistry, Biology, and Clinical ApplicationsFrom EverandTelomerases: Chemistry, Biology, and Clinical ApplicationsNeal F. LueNo ratings yet
- Penetration Tests According To The ISO 11140Document41 pagesPenetration Tests According To The ISO 11140masthan6yNo ratings yet
- ABC PoemDocument3 pagesABC Poemmasthan6yNo ratings yet
- Audit Observation PDocument23 pagesAudit Observation Pmasthan6y100% (1)
- Stock Filters All NewDocument8 pagesStock Filters All Newmasthan6yNo ratings yet
- Purification of MethanolDocument2 pagesPurification of Methanolmasthan6yNo ratings yet
- NEW NEW NEW NEW NEW: Amlodipine BesylateDocument4 pagesNEW NEW NEW NEW NEW: Amlodipine Besylatemasthan6yNo ratings yet
- Daher PromotionDocument1 pageDaher Promotionmasthan6yNo ratings yet
- Date: 01 December 2014 To: Mr. Amjad Ganma Sr. Quality Unit Manager Thru: The Proper Channel Sub: Housing AllowanceDocument1 pageDate: 01 December 2014 To: Mr. Amjad Ganma Sr. Quality Unit Manager Thru: The Proper Channel Sub: Housing Allowancemasthan6yNo ratings yet
- Yakub AnwarDocument3 pagesYakub Anwarmasthan6yNo ratings yet
- Parts of Sterile GarmentDocument1 pageParts of Sterile Garmentmasthan6yNo ratings yet
- Points About WaseelaDocument11 pagesPoints About Waseelamasthan6yNo ratings yet
- Gene Tech Essay PDFDocument4 pagesGene Tech Essay PDFdev_nakomNo ratings yet
- POGIL Model I Mutations PracticeDocument6 pagesPOGIL Model I Mutations PracticeJohana RojasNo ratings yet
- Chapter 17Document15 pagesChapter 17nfnf otupyooorefnNo ratings yet
- Rna PDFDocument49 pagesRna PDFbangbro93_30900715No ratings yet
- Manual BitchesDocument26 pagesManual Bitchessomething1418No ratings yet
- DNA NotesDocument26 pagesDNA NotesMuli Maroshi100% (1)
- Genetic TechnologyDocument36 pagesGenetic TechnologyAnggraeni Kusuma Wardani100% (1)
- From Gene To Protein: How Genes WorkDocument58 pagesFrom Gene To Protein: How Genes WorkGabe GallagherNo ratings yet
- Las and RHL RegulationDocument6 pagesLas and RHL RegulationShouvik BasuNo ratings yet
- Molecular Basis of InheritanceDocument33 pagesMolecular Basis of InheritanceJabcv OahdjNo ratings yet
- Steps of DNA ReplicationDocument5 pagesSteps of DNA Replicationatul mishra100% (1)
- Enhanced Foreign Protein Accumulation in Nicotiana Benthamiana Leaves Co-Infiltrated With A TMV Vector and Plant Cell Cycle Regulator GenesDocument7 pagesEnhanced Foreign Protein Accumulation in Nicotiana Benthamiana Leaves Co-Infiltrated With A TMV Vector and Plant Cell Cycle Regulator Geneszhaoyue12112001No ratings yet
- NCBI Blast - M22-15059-14-MU-2 - 3FDocument3 pagesNCBI Blast - M22-15059-14-MU-2 - 3FCarlitah CortinaNo ratings yet
- Gene LibrariesDocument14 pagesGene Librariessharmamaddy32No ratings yet
- Biotechnology Advances: Pavel Krenek, Olga Samajova, Ivan Luptovciak, Anna Doskocilova, George Komis, Jozef SamajDocument19 pagesBiotechnology Advances: Pavel Krenek, Olga Samajova, Ivan Luptovciak, Anna Doskocilova, George Komis, Jozef SamajAndrés PinzónNo ratings yet
- Bio1A03 Practice Test 2Document11 pagesBio1A03 Practice Test 2cbox4273No ratings yet
- Bm7 RemodellingDocument105 pagesBm7 RemodellingabdullahshiddiqadamNo ratings yet
- BTN 315 Exam Prep ch1 4Document3 pagesBTN 315 Exam Prep ch1 4Corbyn JasonNo ratings yet
- Machine Learning For Designing Next-Generation mRNA TherapeuticsDocument11 pagesMachine Learning For Designing Next-Generation mRNA TherapeuticsJohnNo ratings yet
- Genetics - PYQs and Concept Maps-AKDocument92 pagesGenetics - PYQs and Concept Maps-AKSaiNagaVignesh BandhamNo ratings yet
- Chapter 21-Transgenic Animals: Methodology and ApplicationsDocument24 pagesChapter 21-Transgenic Animals: Methodology and ApplicationsGandis IndahNo ratings yet
- 1.5 Presentasi Ekson IntronDocument30 pages1.5 Presentasi Ekson IntronNessa Muthia RNo ratings yet
- Bicoid Is A: Operator DNA TranscriptionDocument3 pagesBicoid Is A: Operator DNA TranscriptionNiño Matthew Fetalvero RiojaNo ratings yet
- RNA and Protein Synthesis Problem SetDocument6 pagesRNA and Protein Synthesis Problem Setpalms thatshatterNo ratings yet
- 6Document17 pages6AmaniNo ratings yet
- True or FalseDocument4 pagesTrue or Falsetaya guyNo ratings yet
- Plasmids 101 Ebook 3rd Ed FinalDocument193 pagesPlasmids 101 Ebook 3rd Ed FinalManuel CamachoNo ratings yet
- Polymerase Chain Reaction (PCR)Document17 pagesPolymerase Chain Reaction (PCR)pushpavalliNo ratings yet



























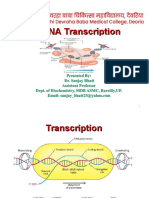










































































Download as ppt, pdf, or txt
You might also like
- Human Molecular Genetics, 4th Edition PDFDocument704 pagesHuman Molecular Genetics, 4th Edition PDFLi Luo100% (5)
- Pro TranscriptionDocument20 pagesPro TranscriptionnitralekhaNo ratings yet
- Methanol Production and UseDocument403 pagesMethanol Production and UseEvin Joy92% (13)
- The Central Dogma of Molecular BiologyDocument4 pagesThe Central Dogma of Molecular BiologyAlvin Fruelda FaaNo ratings yet
- Transcription and RegulationDocument40 pagesTranscription and RegulationRUDRANSH PUJARINo ratings yet
- TranscriptionDocument23 pagesTranscriptionareen fakhouryNo ratings yet
- TranscriptionDocument10 pagesTranscriptionHardik ManekNo ratings yet
- 25 LN RNA Organization and Transcription CNRAADocument55 pages25 LN RNA Organization and Transcription CNRAADakshitha DharmakeerthiNo ratings yet
- G. Gene Expression and Protein SynthesisDocument50 pagesG. Gene Expression and Protein SynthesisMary Rose Bobis VicenteNo ratings yet
- Lecture On Transcription and TranslationDocument47 pagesLecture On Transcription and TranslationAnna Beatrice BautistaNo ratings yet
- DNA Transcription (Part-1) : DR - Ahmed Salim Mohammed PH.D Molecular MicrobiologyDocument49 pagesDNA Transcription (Part-1) : DR - Ahmed Salim Mohammed PH.D Molecular MicrobiologyDrAhmedSalimNo ratings yet
- Webb - DNA 1Document33 pagesWebb - DNA 1monday125No ratings yet
- Gene Expression: Vikrant Joshi F.Y. Biotechnology 6519 Khalsa College AmritsarDocument31 pagesGene Expression: Vikrant Joshi F.Y. Biotechnology 6519 Khalsa College Amritsarvikrant4018No ratings yet
- Topic 10 Transcription Lecture NotesDocument33 pagesTopic 10 Transcription Lecture NoteskambulukatambilaiNo ratings yet
- Nucleic Acid Metabolism TAUDocument38 pagesNucleic Acid Metabolism TAUFaatimah EssaNo ratings yet
- Regulation of Gene Expression in Eukaryotes: Presented by Quratulain (19-Arid-1403) Asadullah (19-Arid-1379)Document32 pagesRegulation of Gene Expression in Eukaryotes: Presented by Quratulain (19-Arid-1403) Asadullah (19-Arid-1379)Beauty LiciousNo ratings yet
- Kuliah 8. Transcription (Oke)Document43 pagesKuliah 8. Transcription (Oke)Sinta DosiNo ratings yet
- 2 TranscriptionDocument51 pages2 Transcriptionmaksumrabbi rumc1999No ratings yet
- Lecture 2Document33 pagesLecture 2SUNDAS FATIMANo ratings yet
- Molecular Biology: TranscriptionDocument101 pagesMolecular Biology: TranscriptionchintyaNo ratings yet
- Central Dogma of Molecular BiologyDocument6 pagesCentral Dogma of Molecular BiologyNiar Ar-RahmahNo ratings yet
- Transcription and TranslationDocument46 pagesTranscription and TranslationEsther SparksNo ratings yet
- Transcription in ProkaryotesDocument16 pagesTranscription in ProkaryotesAditya Kanwal100% (2)
- Transcription and Post-Transcriptional Processing Translation and Post-Translational Protein ProcessingDocument79 pagesTranscription and Post-Transcriptional Processing Translation and Post-Translational Protein ProcessingBcher ChkirNo ratings yet
- RNA Synthesis and ProcessingDocument17 pagesRNA Synthesis and ProcessingInnocent Clifford MaranduNo ratings yet
- Protein SynthesisDocument60 pagesProtein Synthesislucky mbaselaNo ratings yet
- Molecular Biology: TranscriptionDocument101 pagesMolecular Biology: TranscriptiontantyNo ratings yet
- TranscriptionDocument61 pagesTranscriptiondeepak mauryaNo ratings yet
- TranscriptionDocument56 pagesTranscriptionVipin100% (8)
- Transcription: DR Vaishali Dhat Professor, Department of Biochemistry DR DypmcDocument25 pagesTranscription: DR Vaishali Dhat Professor, Department of Biochemistry DR Dypmcnishkarsh chauhanNo ratings yet
- Lecture 11 TranscriptionDocument21 pagesLecture 11 TranscriptionAditi SharmaNo ratings yet
- Transciption and TranslationDocument31 pagesTransciption and TranslationDuduetsang MosalakataneNo ratings yet
- 6 Lecture-TranslationDocument25 pages6 Lecture-Translationmoazzanrana678No ratings yet
- Transcription: From DNA To RNADocument74 pagesTranscription: From DNA To RNAmd habibur rahmanNo ratings yet
- UntitledDocument45 pagesUntitledjemNo ratings yet
- TRANSKRIPSIDocument19 pagesTRANSKRIPSIZakisyarifudintaqiyNo ratings yet
- Sintesis DNADocument32 pagesSintesis DNAZhafran DarwisNo ratings yet
- Genetik MikrobDocument33 pagesGenetik MikrobekasukmawatyNo ratings yet
- Rna Biosynthesis (Transicription)Document33 pagesRna Biosynthesis (Transicription)Alaa AlmajedNo ratings yet
- Inhibition of DNA Replication and TranscriptionDocument111 pagesInhibition of DNA Replication and TranscriptionPranav Kumar PrabhakarNo ratings yet
- Gene ExpressionDocument64 pagesGene ExpressiondesyNo ratings yet
- Gene ExpressionDocument72 pagesGene ExpressionMika ForwardNo ratings yet
- Science FairDocument41 pagesScience FairSafanaNo ratings yet
- Lesson 9Document66 pagesLesson 9Irica Mae CiervoNo ratings yet
- 106 Ceng w10Document62 pages106 Ceng w10mertNo ratings yet
- Chapter 5 Protein SynthesisDocument26 pagesChapter 5 Protein SynthesisT MokshithaNo ratings yet
- 22 (W13) TranscriptionDocument38 pages22 (W13) TranscriptionMUHAMMAD FAHMI MOHD ZAMZANINo ratings yet
- Transcription 2Document66 pagesTranscription 2Safe BoxNo ratings yet
- Ekspresi GenDocument63 pagesEkspresi GenIndraPramanaIdaBagusNo ratings yet
- PROCESS of TRANSKRIPSI, TRANSLASI, REPLIKASI DNA2Document27 pagesPROCESS of TRANSKRIPSI, TRANSLASI, REPLIKASI DNA2istatinNo ratings yet
- TUTORIAL: DNA BIOLOGY and TECHNOLOGY 1. Describe The Biochemical CompositionDocument6 pagesTUTORIAL: DNA BIOLOGY and TECHNOLOGY 1. Describe The Biochemical Compositionaesha89No ratings yet
- Biochemistry: RNA Synthesis and ProcessingDocument54 pagesBiochemistry: RNA Synthesis and ProcessingAqsa YaminNo ratings yet
- Transcription ProkaryoticDocument30 pagesTranscription ProkaryoticDibya Jyoti ParidaNo ratings yet
- RNA Processing or Post Transcriptional Modifications.Document23 pagesRNA Processing or Post Transcriptional Modifications.saeed313bbtNo ratings yet
- DNA-Dependent Synthesis of RNA: (Transcription)Document10 pagesDNA-Dependent Synthesis of RNA: (Transcription)Shivam SharmaNo ratings yet
- TranscriptionDocument34 pagesTranscriptiondrhydrogenNo ratings yet
- Lecture 5 Diagnostic GenomicsDocument33 pagesLecture 5 Diagnostic Genomicssales zfNo ratings yet
- Lect 4 Prokaryotic Gene StructureDocument45 pagesLect 4 Prokaryotic Gene StructureElias LibayNo ratings yet
- TransDocument29 pagesTransDrLakshmi Priya ThyagarajanNo ratings yet
- Chapter 6 SummaryDocument28 pagesChapter 6 SummaryCharlotteNo ratings yet
- Telomerases: Chemistry, Biology, and Clinical ApplicationsFrom EverandTelomerases: Chemistry, Biology, and Clinical ApplicationsNeal F. LueNo ratings yet
- Penetration Tests According To The ISO 11140Document41 pagesPenetration Tests According To The ISO 11140masthan6yNo ratings yet
- ABC PoemDocument3 pagesABC Poemmasthan6yNo ratings yet
- Audit Observation PDocument23 pagesAudit Observation Pmasthan6y100% (1)
- Stock Filters All NewDocument8 pagesStock Filters All Newmasthan6yNo ratings yet
- Purification of MethanolDocument2 pagesPurification of Methanolmasthan6yNo ratings yet
- NEW NEW NEW NEW NEW: Amlodipine BesylateDocument4 pagesNEW NEW NEW NEW NEW: Amlodipine Besylatemasthan6yNo ratings yet
- Daher PromotionDocument1 pageDaher Promotionmasthan6yNo ratings yet
- Date: 01 December 2014 To: Mr. Amjad Ganma Sr. Quality Unit Manager Thru: The Proper Channel Sub: Housing AllowanceDocument1 pageDate: 01 December 2014 To: Mr. Amjad Ganma Sr. Quality Unit Manager Thru: The Proper Channel Sub: Housing Allowancemasthan6yNo ratings yet
- Yakub AnwarDocument3 pagesYakub Anwarmasthan6yNo ratings yet
- Parts of Sterile GarmentDocument1 pageParts of Sterile Garmentmasthan6yNo ratings yet
- Points About WaseelaDocument11 pagesPoints About Waseelamasthan6yNo ratings yet
- Gene Tech Essay PDFDocument4 pagesGene Tech Essay PDFdev_nakomNo ratings yet
- POGIL Model I Mutations PracticeDocument6 pagesPOGIL Model I Mutations PracticeJohana RojasNo ratings yet
- Chapter 17Document15 pagesChapter 17nfnf otupyooorefnNo ratings yet
- Rna PDFDocument49 pagesRna PDFbangbro93_30900715No ratings yet
- Manual BitchesDocument26 pagesManual Bitchessomething1418No ratings yet
- DNA NotesDocument26 pagesDNA NotesMuli Maroshi100% (1)
- Genetic TechnologyDocument36 pagesGenetic TechnologyAnggraeni Kusuma Wardani100% (1)
- From Gene To Protein: How Genes WorkDocument58 pagesFrom Gene To Protein: How Genes WorkGabe GallagherNo ratings yet
- Las and RHL RegulationDocument6 pagesLas and RHL RegulationShouvik BasuNo ratings yet
- Molecular Basis of InheritanceDocument33 pagesMolecular Basis of InheritanceJabcv OahdjNo ratings yet
- Steps of DNA ReplicationDocument5 pagesSteps of DNA Replicationatul mishra100% (1)
- Enhanced Foreign Protein Accumulation in Nicotiana Benthamiana Leaves Co-Infiltrated With A TMV Vector and Plant Cell Cycle Regulator GenesDocument7 pagesEnhanced Foreign Protein Accumulation in Nicotiana Benthamiana Leaves Co-Infiltrated With A TMV Vector and Plant Cell Cycle Regulator Geneszhaoyue12112001No ratings yet
- NCBI Blast - M22-15059-14-MU-2 - 3FDocument3 pagesNCBI Blast - M22-15059-14-MU-2 - 3FCarlitah CortinaNo ratings yet
- Gene LibrariesDocument14 pagesGene Librariessharmamaddy32No ratings yet
- Biotechnology Advances: Pavel Krenek, Olga Samajova, Ivan Luptovciak, Anna Doskocilova, George Komis, Jozef SamajDocument19 pagesBiotechnology Advances: Pavel Krenek, Olga Samajova, Ivan Luptovciak, Anna Doskocilova, George Komis, Jozef SamajAndrés PinzónNo ratings yet
- Bio1A03 Practice Test 2Document11 pagesBio1A03 Practice Test 2cbox4273No ratings yet
- Bm7 RemodellingDocument105 pagesBm7 RemodellingabdullahshiddiqadamNo ratings yet
- BTN 315 Exam Prep ch1 4Document3 pagesBTN 315 Exam Prep ch1 4Corbyn JasonNo ratings yet
- Machine Learning For Designing Next-Generation mRNA TherapeuticsDocument11 pagesMachine Learning For Designing Next-Generation mRNA TherapeuticsJohnNo ratings yet
- Genetics - PYQs and Concept Maps-AKDocument92 pagesGenetics - PYQs and Concept Maps-AKSaiNagaVignesh BandhamNo ratings yet
- Chapter 21-Transgenic Animals: Methodology and ApplicationsDocument24 pagesChapter 21-Transgenic Animals: Methodology and ApplicationsGandis IndahNo ratings yet
- 1.5 Presentasi Ekson IntronDocument30 pages1.5 Presentasi Ekson IntronNessa Muthia RNo ratings yet
- Bicoid Is A: Operator DNA TranscriptionDocument3 pagesBicoid Is A: Operator DNA TranscriptionNiño Matthew Fetalvero RiojaNo ratings yet
- RNA and Protein Synthesis Problem SetDocument6 pagesRNA and Protein Synthesis Problem Setpalms thatshatterNo ratings yet
- 6Document17 pages6AmaniNo ratings yet
- True or FalseDocument4 pagesTrue or Falsetaya guyNo ratings yet
- Plasmids 101 Ebook 3rd Ed FinalDocument193 pagesPlasmids 101 Ebook 3rd Ed FinalManuel CamachoNo ratings yet
- Polymerase Chain Reaction (PCR)Document17 pagesPolymerase Chain Reaction (PCR)pushpavalliNo ratings yet