Download as pptx, pdf, or txt
You might also like
- Personal Training Level 3 ManualDocument358 pagesPersonal Training Level 3 ManualAATdale94% (18)
- TestBank Norris Porths Essentials Pathophysiology 5e 2019Document505 pagesTestBank Norris Porths Essentials Pathophysiology 5e 2019Cindy Perez50% (2)
- Home Production of Quality Meats and SausagesDocument566 pagesHome Production of Quality Meats and SausagesThalyta Cruxz100% (14)
- 2.membrane Physiology, Nerve and MuscleDocument6 pages2.membrane Physiology, Nerve and MuscleCLEMENT100% (1)
- Muscle Structure and FunctionsDocument59 pagesMuscle Structure and FunctionsThopu Umamaheswari100% (1)
- The Nervous SystemDocument71 pagesThe Nervous SystemDennis Limosnero MayorNo ratings yet
- Chapter 27 GuytonDocument6 pagesChapter 27 GuytonGabriella ChafrinaNo ratings yet
- Threshold and RecruitmentDocument16 pagesThreshold and RecruitmentShamaine Anne SalvadorNo ratings yet
- Chapter - 13 - Cardiovascular - Responses - Exercise Physiology For Health Fitness and PerformanceDocument32 pagesChapter - 13 - Cardiovascular - Responses - Exercise Physiology For Health Fitness and Performanceec16043No ratings yet
- Water LoseDocument29 pagesWater LoseJAKLIN EMPOLNo ratings yet
- Basic Clinical Massage TherapyDocument654 pagesBasic Clinical Massage Therapykkism100% (11)
- Lecture 1 Physiology of Skeletal Muscle by Dr. RoomiDocument37 pagesLecture 1 Physiology of Skeletal Muscle by Dr. RoomiMudassar Roomi100% (1)
- Muscle PhysiologyDocument49 pagesMuscle PhysiologyNauval Zilal FananyNo ratings yet
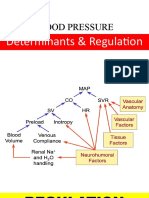
- Short and Long Term Regulation and Determinants of BPDocument90 pagesShort and Long Term Regulation and Determinants of BPDan Tristam MicabaloNo ratings yet
- Chemical Coordination and IntegrationDocument6 pagesChemical Coordination and IntegrationSharanya GhoshNo ratings yet
- MHC I Proteins, Which Present Antigens To Cytotoxic T Cells, (2) MHC II Proteins, Which Present Antigens T Helper CellsDocument25 pagesMHC I Proteins, Which Present Antigens To Cytotoxic T Cells, (2) MHC II Proteins, Which Present Antigens T Helper CellsMudassar Roomi100% (1)
- 10 Muscular ContractionDocument29 pages10 Muscular ContractionMark Piedad MillanoNo ratings yet
- 6-8 Thrombolitics Antiplatelet AnticoagulantDocument36 pages6-8 Thrombolitics Antiplatelet AnticoagulantFatima ZahraNo ratings yet
- Bio Mechanics Lecture NotesDocument53 pagesBio Mechanics Lecture NotesscdubalNo ratings yet
- Blood, Vessels and Pressure Phys LectDocument23 pagesBlood, Vessels and Pressure Phys LectVivek ChaudharyNo ratings yet
- Renal Physiology: Done byDocument28 pagesRenal Physiology: Done byJanine Marie Kathleen Uy-CuanangNo ratings yet
- Introduction To ANS PharmacologyDocument34 pagesIntroduction To ANS PharmacologySebontu HasenNo ratings yet
- Pharmacology Trans ANS Drugs IDocument20 pagesPharmacology Trans ANS Drugs IPrincess Mara DuranNo ratings yet
- 1st Lec On Heart Physiology by Dr. RoomiDocument13 pages1st Lec On Heart Physiology by Dr. RoomiMudassar Roomi100% (1)
- Week12 13 RENAL PhysiologyDocument41 pagesWeek12 13 RENAL Physiologyirwan kastellaNo ratings yet
- Medical Mnemonic BookletDocument14 pagesMedical Mnemonic Bookletahmed abd elazizNo ratings yet
- Pharmacology of Autonomic Nervous SystemDocument106 pagesPharmacology of Autonomic Nervous Systemoscar3spurgeonNo ratings yet
- Pharmacology Antibiotics Table FINALDocument3 pagesPharmacology Antibiotics Table FINALAndre AndreeaNo ratings yet
- 1 The Electrical Activity of The HeartDocument2 pages1 The Electrical Activity of The HeartRickertsNo ratings yet
- Autonomic SystemDocument29 pagesAutonomic Systemsanaya100% (1)
- Unit 8 Learning Guide Name: Brandon Au-Young: InstructionsDocument25 pagesUnit 8 Learning Guide Name: Brandon Au-Young: InstructionsBrandonNo ratings yet
- Basics On NeurophysiologyDocument12 pagesBasics On NeurophysiologyRausche Anne Blaser Sausa100% (1)
- Skeletal Muscle: Lecturer: DR HERMAN MULIJADI, MS. SP KPDocument43 pagesSkeletal Muscle: Lecturer: DR HERMAN MULIJADI, MS. SP KPAriana KimNo ratings yet
- Cholinergic and AnticholinergicDocument77 pagesCholinergic and Anticholinergicsweta sumanNo ratings yet
- 10.2 Mechanism of Muscle ContractionDocument38 pages10.2 Mechanism of Muscle ContractionDaksha yashaNo ratings yet
- 2 - Renal PhysiologyDocument8 pages2 - Renal PhysiologyKunware TropaNo ratings yet
- Physiology: Guevara, Henaku-Larbi, Inciong, JutizDocument3 pagesPhysiology: Guevara, Henaku-Larbi, Inciong, JutizMavic VillanuevaNo ratings yet
- Functional Human Physiology: For The Exercise and Sport Sciences The Cardiovascular System: Cardiac FunctionDocument186 pagesFunctional Human Physiology: For The Exercise and Sport Sciences The Cardiovascular System: Cardiac FunctionBery Agana F. PurbaNo ratings yet
- Cardio Day 3 TemplateDocument24 pagesCardio Day 3 TemplateMikeNo ratings yet
- Blood: 1. Medullary Cavities of Long Bones 2. Spongiosa of Vertebral Bodies, Ribs, Sternum 3. Flat Bones of The PelvisDocument2 pagesBlood: 1. Medullary Cavities of Long Bones 2. Spongiosa of Vertebral Bodies, Ribs, Sternum 3. Flat Bones of The PelvisKrizza Mariz Bautista PolaronNo ratings yet
- Chapt 05Document83 pagesChapt 05emmielyaNo ratings yet
- Important SEQs Heart Physiology by Dr. RoomiDocument3 pagesImportant SEQs Heart Physiology by Dr. RoomiMudassar Roomi100% (2)
- NERVE & MUSCLE 45 Qs With Their Answer Keys by Dr. RoomiDocument54 pagesNERVE & MUSCLE 45 Qs With Their Answer Keys by Dr. RoomiMudassar Roomi100% (1)
- Immunology Rheumatology NotesDocument178 pagesImmunology Rheumatology NotesameenaalqasimNo ratings yet
- Cell CommunicationDocument45 pagesCell CommunicationWilson HindartoNo ratings yet
- Nervous System: (Ex. Acetylcholine, Norepinephrine)Document65 pagesNervous System: (Ex. Acetylcholine, Norepinephrine)Karl Nielo BartolomeNo ratings yet
- Anatomy Lecture 19,20,21: 1 Hour 04,06,08-11-2013Document64 pagesAnatomy Lecture 19,20,21: 1 Hour 04,06,08-11-2013Batot EvaNo ratings yet
- Introduction To HormonesDocument41 pagesIntroduction To Hormonesنورالهدى حسام عليNo ratings yet
- Endocrine Notes - All in One FileDocument182 pagesEndocrine Notes - All in One FilekjNo ratings yet
- PARASYMPATHETIC GANGLION in Head and NeckDocument53 pagesPARASYMPATHETIC GANGLION in Head and NeckDEEP SABNo ratings yet
- Renal Physiology and Fluid BalanceDocument71 pagesRenal Physiology and Fluid BalanceparthNo ratings yet
- Cardiovascular Pharmacology PDFDocument85 pagesCardiovascular Pharmacology PDFAhmed Shihab AhmedNo ratings yet
- Anatomy Atlas Neuroanatomy Fawzy GaballahDocument228 pagesAnatomy Atlas Neuroanatomy Fawzy GaballahNicolas Octavio100% (1)
- Anti Asthmatic DrugsDocument8 pagesAnti Asthmatic DrugsNavjot BrarNo ratings yet
- Key of SEQs Heart, Nerve and Muscle Revision TestDocument11 pagesKey of SEQs Heart, Nerve and Muscle Revision TestMudassar Roomi100% (2)
- OPP Lecture 1Document7 pagesOPP Lecture 1Lenny CostantiniNo ratings yet
- Autonomic Nervous System PharmacologyDocument97 pagesAutonomic Nervous System PharmacologyKarun Kumar Shingari100% (1)
- Renal PhysiologyDocument75 pagesRenal PhysiologyAmanuel Maru100% (1)
- Physiology Cardiovascular Physiology NotesDocument13 pagesPhysiology Cardiovascular Physiology Noteshedayatamin44No ratings yet
- 8 - Functional Organization of The Nervous System Moyes 3rd Ed 2018 PDFDocument71 pages8 - Functional Organization of The Nervous System Moyes 3rd Ed 2018 PDFLiane LomioNo ratings yet
- Contraction of Skeletal MuscleDocument61 pagesContraction of Skeletal MusclendorokemewisdomNo ratings yet
- Chapter 5Document25 pagesChapter 5أمال داودNo ratings yet
- Skeletal MusclesDocument7 pagesSkeletal MusclesAhmed OudahNo ratings yet
- Muscle Physiology in Orthodontics / Orthodontic Courses by Indian Dental AcademyDocument50 pagesMuscle Physiology in Orthodontics / Orthodontic Courses by Indian Dental Academyindian dental academyNo ratings yet
- Excitation Contraction Coupling: Nandini GoyalDocument32 pagesExcitation Contraction Coupling: Nandini GoyalNandini GoyalNo ratings yet
- Farmakokinetik & Farmakodinamik Ujian: Pemimbing Andri Tilaqza, S.Farm, M.Farm, AptvDocument35 pagesFarmakokinetik & Farmakodinamik Ujian: Pemimbing Andri Tilaqza, S.Farm, M.Farm, AptvMakmunNawilNo ratings yet
- Medication Overuse HeadacheDocument5 pagesMedication Overuse HeadacheMakmunNawilNo ratings yet
- Case Report Measles-Mumps-Rubella Vaccination Induced Thrombocytopenia: A Case Report and Review of The LiteratureDocument11 pagesCase Report Measles-Mumps-Rubella Vaccination Induced Thrombocytopenia: A Case Report and Review of The LiteratureMakmunNawilNo ratings yet
- Cushing's Syndrome: Oleh: Kelompok VDocument15 pagesCushing's Syndrome: Oleh: Kelompok VMakmunNawilNo ratings yet
- RubelaDocument5 pagesRubelaMakmunNawilNo ratings yet
- Infective Endocarditis ESC 09Document27 pagesInfective Endocarditis ESC 09MakmunNawilNo ratings yet
- Submodule 2.5 - 2.6Document14 pagesSubmodule 2.5 - 2.6Zeejiah NelleNo ratings yet
- Module in Meat Science and Technology CO1Document28 pagesModule in Meat Science and Technology CO1NALANG FEBREA MAE D.No ratings yet
- Chapter 8 - Musculoskeletal SystemDocument9 pagesChapter 8 - Musculoskeletal SystemIbby HooriyaNo ratings yet
- Chemistry in The Kitchen: The Chemistry of Flesh Foods II: Peter BaylissDocument6 pagesChemistry in The Kitchen: The Chemistry of Flesh Foods II: Peter BaylissMajo ArguelloNo ratings yet
- Nerve Muscle PhysiologyDocument18 pagesNerve Muscle PhysiologyDr P N N Reddy100% (2)
- 2nd Year BioDocument2 pages2nd Year BioJaveria ZaidiNo ratings yet
- MuscularDocument61 pagesMuscularFranz DiazNo ratings yet
- Anatomy ReviewDocument6 pagesAnatomy ReviewKwai Fong ChanNo ratings yet
- Muscle Tissue NotesDocument11 pagesMuscle Tissue NotesAubrey Unique EvangelistaNo ratings yet
- Clinical Case: Anatomy and PhysiologyDocument73 pagesClinical Case: Anatomy and PhysiologyGeorge KoNo ratings yet
- Biol 273 NotesDocument46 pagesBiol 273 NotesDivyaNo ratings yet
- Lehninger PPT ch05Document62 pagesLehninger PPT ch05MahdeeHaqueSyed100% (1)
- Human Anatomy,: First Edition Mckinley & O'LoughlinDocument77 pagesHuman Anatomy,: First Edition Mckinley & O'LoughlinHarman BainzNo ratings yet
- Lehninger - 7e - PPT - ch05 - Final (Autosaved)Document53 pagesLehninger - 7e - PPT - ch05 - Final (Autosaved)Chance SwansonNo ratings yet
- Chapter 7 - Muscuar SystemDocument17 pagesChapter 7 - Muscuar SystemM GarciaNo ratings yet
- Make-Up of The Human BodyDocument7 pagesMake-Up of The Human BodyMariam GamosNo ratings yet
- AHS 131 Lecture 4 MusclesDocument60 pagesAHS 131 Lecture 4 MusclesBikash MainaliNo ratings yet
- Zoology PPT Muscular SystemDocument68 pagesZoology PPT Muscular SystemDian O. Velasquez100% (2)
- Case Study Wearing On Her NervesDocument42 pagesCase Study Wearing On Her Nervesapi-460556390No ratings yet
- The Muscular SystemDocument18 pagesThe Muscular SystemSilent GlitchNo ratings yet
- Tortora Outline Chapter 10 For PPDocument12 pagesTortora Outline Chapter 10 For PPTasha BradyNo ratings yet
- Ranking of Beef Muscles For Tenderness PDFDocument6 pagesRanking of Beef Muscles For Tenderness PDFNaomi KangNo ratings yet
- Classification of MusclesDocument50 pagesClassification of MusclesNoor HaleemNo ratings yet
- Lectures 8-10 NotesDocument15 pagesLectures 8-10 Notesapi-3721438No ratings yet