Download as ppt, pdf, or txt
You might also like
- Mock Exam 1Document34 pagesMock Exam 1Althea Karmylle M. BonitaNo ratings yet
- LECTURE 12 TranslationDocument17 pagesLECTURE 12 TranslationAditi SharmaNo ratings yet
- Translation 190603145457Document30 pagesTranslation 190603145457soniaazizNo ratings yet
- 8d) Translation Summary - 9744 - 2017Document1 page8d) Translation Summary - 9744 - 2017saguNo ratings yet
- Translation - IDocument35 pagesTranslation - ISahil RanaNo ratings yet
- 8d) Translation Summary - 9744 - 2018Document1 page8d) Translation Summary - 9744 - 20182022 EMMA WEN XUAN HANSONNo ratings yet
- The Translation: Translation Is A Process by Which The Genetic Code Contained Within An mRNADocument5 pagesThe Translation: Translation Is A Process by Which The Genetic Code Contained Within An mRNAShahriar ShamimNo ratings yet
- The Birth and Death of Proteins: Some Key ConceptsDocument2 pagesThe Birth and Death of Proteins: Some Key ConceptsLuke ShantiNo ratings yet
- 2022-05-15 L8 - Protein SynthesisDocument64 pages2022-05-15 L8 - Protein SynthesisTamara ElyasNo ratings yet
- Translation: Mrs. Ofelia Solano SaludarDocument28 pagesTranslation: Mrs. Ofelia Solano SaludarmskikiNo ratings yet
- Translation 1Document68 pagesTranslation 1Disha GuptaNo ratings yet
- 13 Miller Chap 4b LectureDocument21 pages13 Miller Chap 4b LectureGerone Tolentino AtienzaNo ratings yet
- TranslationDocument90 pagesTranslationspitzmark2030No ratings yet
- Advanced Chemistryprize2009 PDFDocument24 pagesAdvanced Chemistryprize2009 PDFJeremy GordonNo ratings yet
- Lec. 6Document10 pagesLec. 6Dr. Mohamed ShamsNo ratings yet
- MB Chapter 6 TranslationDocument31 pagesMB Chapter 6 TranslationMustee TeferaNo ratings yet
- 12TrlAMO LicyayoDocument9 pages12TrlAMO LicyayoMohamidin MamalapatNo ratings yet
- Translation in Prokaryotes: NEET 2020Document1 pageTranslation in Prokaryotes: NEET 2020ADIKKI ANOOHYANo ratings yet
- Central Dogma: Translation NotesDocument9 pagesCentral Dogma: Translation NotesRomel Greg TulangNo ratings yet
- MBII - L23 - Translation 2Document9 pagesMBII - L23 - Translation 2Miles NsgNo ratings yet
- TranslationDocument51 pagesTranslationAleena MustafaNo ratings yet
- Agus Limanto Faculty of Medicine UkridaDocument26 pagesAgus Limanto Faculty of Medicine UkridaMrs GeekNo ratings yet
- Translation: The Last Stage of Central DogmaDocument10 pagesTranslation: The Last Stage of Central DogmaChandu Kishore KalloniNo ratings yet
- Lecture 3Document6 pagesLecture 3tiaria.wilson24No ratings yet
- L5-6 Translation in ProkaryotesDocument17 pagesL5-6 Translation in ProkaryotesArchana BhartiNo ratings yet
- Otot 2Document106 pagesOtot 2anita parwatiNo ratings yet
- Protien Synthesis Elongation & TerminationDocument14 pagesProtien Synthesis Elongation & TerminationvesascNo ratings yet
- The Mechanism of TranslationDocument68 pagesThe Mechanism of TranslationRama KishoreNo ratings yet
- Translation 30th March 2009Document16 pagesTranslation 30th March 2009api-26794308No ratings yet
- Translation in ProkaryotesDocument7 pagesTranslation in ProkaryotesAkachukwu ObunikeNo ratings yet
- 6 of 7 Molecular BiologyDocument48 pages6 of 7 Molecular BiologyAzhar MehmoodNo ratings yet
- TRANSLATIONDocument16 pagesTRANSLATIONkhushboo bhardwajNo ratings yet
- TranslationDocument1 pageTranslationGianne AbcedeNo ratings yet
- Translation AnoverviewDocument32 pagesTranslation AnoverviewshitalchandrasitNo ratings yet
- TranslationDocument27 pagesTranslationzzzNo ratings yet
- Translation PresentationDocument32 pagesTranslation PresentationAchrya KailashNo ratings yet
- Central Dogma: Faculty of Medicine, Sriwijaya UniversityDocument51 pagesCentral Dogma: Faculty of Medicine, Sriwijaya UniversityIlsya PertiwiNo ratings yet
- Translation-An Overview: Professor (DR.) Namrata Chhabra Biochemistry For Medics - Lecture Notes WWW - Namrata.coDocument32 pagesTranslation-An Overview: Professor (DR.) Namrata Chhabra Biochemistry For Medics - Lecture Notes WWW - Namrata.coDurga Charan PradhanNo ratings yet
- Translation (Biology)Document22 pagesTranslation (Biology)Manoj JoshiNo ratings yet
- Translation: The Synthesis of Polypeptides Using Mrna TemplatesDocument4 pagesTranslation: The Synthesis of Polypeptides Using Mrna TemplatesMudjaijah draNo ratings yet
- Cell Biology: InstructorDocument17 pagesCell Biology: Instructorahmed mediaNo ratings yet
- The Mechanism of Protein Synthesis: Initiation: Elongation: TerminationDocument16 pagesThe Mechanism of Protein Synthesis: Initiation: Elongation: TerminationAminNo ratings yet
- Random-Splitting of tRNA Transcripts As An Approach For Studying tRNA-protein InteractionsDocument4 pagesRandom-Splitting of tRNA Transcripts As An Approach For Studying tRNA-protein InteractionslemNo ratings yet
- TRANSLATION Class 12Document4 pagesTRANSLATION Class 12hipulkitguptaNo ratings yet
- Bio Short Notes FinalDocument5 pagesBio Short Notes FinalMUHAMMAD HAFIZ FAZRISHAH BIN JEFFRY KIMNo ratings yet
- DNA Translation - Dr. Mohammed OsmanDocument43 pagesDNA Translation - Dr. Mohammed Osmanمعتز محمدNo ratings yet
- Regulasi Ekspresi GenDocument41 pagesRegulasi Ekspresi GenJohn'sSujonoNo ratings yet
- TRANSLATIONDocument19 pagesTRANSLATIONUsha SuthersonNo ratings yet
- Translation and Regulation of Gene ExpressionDocument51 pagesTranslation and Regulation of Gene ExpressionP. Jacksen Sam PaulNo ratings yet
- 10.06 Prokaryotic TranslationDocument4 pages10.06 Prokaryotic TranslationWidya NingsiNo ratings yet
- Translation: M.Prasad Naidu MSC Medical Biochemistry, PH.DDocument33 pagesTranslation: M.Prasad Naidu MSC Medical Biochemistry, PH.DDr. M. Prasad NaiduNo ratings yet
- Ekspresi Gen PDFDocument36 pagesEkspresi Gen PDFainulavidaNo ratings yet
- Chapter 6 GeneticsDocument12 pagesChapter 6 GeneticsAmer ToutonjiNo ratings yet
- TranslationDocument2 pagesTranslationYash VardhanNo ratings yet
- Translation: The Biochemistry Department of SYMCDocument46 pagesTranslation: The Biochemistry Department of SYMCPascal TurksonNo ratings yet
- Transcription &translation: Mol. Biology Lec-4 TranscriptionDocument12 pagesTranscription &translation: Mol. Biology Lec-4 TranscriptionAhmed Ali AssafNo ratings yet
- TranslationDocument12 pagesTranslationHarshit YadavNo ratings yet
- Protein SynthesisDocument59 pagesProtein SynthesisBalew GetaNo ratings yet
- Translation in ProkaryotesDocument25 pagesTranslation in ProkaryotesNaomi Lamtiur0% (1)
- 02 Protein Synthesis Translation 1Document16 pages02 Protein Synthesis Translation 1Dr Hotimah HotimahNo ratings yet
- Marks Distribution SchemeDocument2 pagesMarks Distribution SchemeAdith SaiNo ratings yet
- Kranthi (Minor 3)Document39 pagesKranthi (Minor 3)Adith SaiNo ratings yet
- PY251 A P Anagha NotesDocument136 pagesPY251 A P Anagha NotesAdith SaiNo ratings yet
- Problems 2017Document2 pagesProblems 2017Adith SaiNo ratings yet
- ThoughtsDocument1 pageThoughtsAdith SaiNo ratings yet
- Time-Energy Uncertainty RelationDocument14 pagesTime-Energy Uncertainty RelationAdith SaiNo ratings yet
- Cardiac Cycle: Definition: The Mechanical Events, Occurring in The Heart Within One Beat (From The Beginning of A HeartDocument6 pagesCardiac Cycle: Definition: The Mechanical Events, Occurring in The Heart Within One Beat (From The Beginning of A HeartAdith SaiNo ratings yet
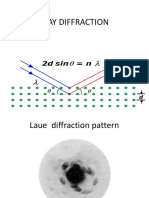
- X Ray DiffractionDocument15 pagesX Ray DiffractionAdith SaiNo ratings yet
- Sociology and Common SenseDocument6 pagesSociology and Common SenseAdith SaiNo ratings yet
- %assignment - 1: Forced - Damped OscillationDocument1 page%assignment - 1: Forced - Damped OscillationAdith SaiNo ratings yet
- Between The Newly Synthesized Strand and The Template StrandDocument52 pagesBetween The Newly Synthesized Strand and The Template StrandAdith SaiNo ratings yet
- DNA Synthesis Occurs During The S Resulting in The Duplication of ChromosomeDocument20 pagesDNA Synthesis Occurs During The S Resulting in The Duplication of ChromosomeAdith SaiNo ratings yet
- Shikimic Acid Pathway: By: Resane Ma. JannelleDocument14 pagesShikimic Acid Pathway: By: Resane Ma. JannelleMA. JANNELLE P. RESANENo ratings yet
- Las Q4 Week-2 GenDocument8 pagesLas Q4 Week-2 GenFranzhean Balais CuachonNo ratings yet
- Yeast Micronutrient Requirements 2017Document4 pagesYeast Micronutrient Requirements 2017Vy NguyenNo ratings yet
- Industrial Wastewater Treatment by AS PDFDocument404 pagesIndustrial Wastewater Treatment by AS PDFGustavo Mayen100% (2)
- Bio Ninja Unit 8Document4 pagesBio Ninja Unit 8shwooby shawoobyNo ratings yet
- Lactic Acidosis ElyaDocument19 pagesLactic Acidosis Elyaapi-303542051No ratings yet
- Learning Module General Biology 1Document15 pagesLearning Module General Biology 1John Paull CuaNo ratings yet
- By The End of This Lecture You Will Be Able To:: Light-Dependent Reactions (I.e., Light Reactions)Document35 pagesBy The End of This Lecture You Will Be Able To:: Light-Dependent Reactions (I.e., Light Reactions)yu90210No ratings yet
- Urea CycleDocument38 pagesUrea CycleAbdimalik Ali100% (1)
- O e Re FH e K e E: Teacher ManualDocument48 pagesO e Re FH e K e E: Teacher Manualpk rNo ratings yet
- Tritest S.R.L.: Buletin de Analize Medicale 36 - 1218Document2 pagesTritest S.R.L.: Buletin de Analize Medicale 36 - 1218CATALINA AVASILCEINo ratings yet
- Digital Unit Plan TemplateDocument5 pagesDigital Unit Plan Templateapi-257521568No ratings yet
- A2 Biology Notes - PP & MS - Energy and Respiration - 2019Document226 pagesA2 Biology Notes - PP & MS - Energy and Respiration - 2019Kulsoom JawedNo ratings yet
- Urine AnalysisDocument3 pagesUrine AnalysisFara Liyana RahimNo ratings yet
- Manuscript NPJDocument24 pagesManuscript NPJDanang RaharjoNo ratings yet
- Deuterium Depletion EbookDocument26 pagesDeuterium Depletion EbookCK_2023No ratings yet
- Characteristics of Living Organisms: All Vertebrates Have A Backbone. There Are 5 Classes of VertebratesDocument4 pagesCharacteristics of Living Organisms: All Vertebrates Have A Backbone. There Are 5 Classes of VertebratesKrish PatelNo ratings yet
- Arcebido, GwynnDocument4 pagesArcebido, GwynnGwynnNo ratings yet
- Cell Theory, MSHGRENDocument6 pagesCell Theory, MSHGREN1301aanyaNo ratings yet
- Cellular Respiration and Fermentation: For Campbell Biology, Ninth EditionDocument37 pagesCellular Respiration and Fermentation: For Campbell Biology, Ninth EditionJovelyn GuerraNo ratings yet
- 92.physiology of Lens and Cataractogenesis... SrisDocument92 pages92.physiology of Lens and Cataractogenesis... SrisSristi ThakurNo ratings yet
- Section XI - Fatty Acid MetabolismDocument3 pagesSection XI - Fatty Acid MetabolismKatharine NervaNo ratings yet
- PhotosynthesisDocument4 pagesPhotosynthesisapi-3729258100% (1)
- William G. Hopkins Plant Biotechnology PDFDocument153 pagesWilliam G. Hopkins Plant Biotechnology PDFcarlos100% (1)
- Amino Acid CatabolismDocument24 pagesAmino Acid CatabolismAnonymous 0ha8TmNo ratings yet
- Gen Bio1 Module 9Document23 pagesGen Bio1 Module 9Anjhiene Camba100% (2)
- Metabolic PathwaysDocument6 pagesMetabolic PathwaysIvanNo ratings yet
- CH 41 Animal NutritionDocument70 pagesCH 41 Animal NutritionHazel EdillorNo ratings yet