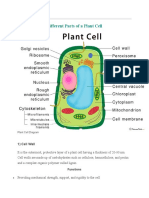
Download as ppt, pdf, or txt
You might also like
- Owners Manual For The Human BodyDocument184 pagesOwners Manual For The Human BodyKika Spacey Ratm100% (2)
- BCH 4107 - Plant BiochemistryDocument13 pagesBCH 4107 - Plant BiochemistryOLUWASEGUN K AfolabiNo ratings yet
- CellDocument61 pagesCellAbdul Rehman100% (1)
- Plant Cells Structure of The Plant CellsDocument10 pagesPlant Cells Structure of The Plant Cellsstudent10100No ratings yet
- Cell Biology Lecture VDocument22 pagesCell Biology Lecture Vbusachala517No ratings yet
- 2 Cell WallDocument9 pages2 Cell WallMuhammad TaimoorNo ratings yet
- Carbohydrate: The Cell WallDocument3 pagesCarbohydrate: The Cell WallFarah Fadhilah RosyadiNo ratings yet
- Eukaryotic Cell StructureDocument40 pagesEukaryotic Cell StructureJevaughn SmithNo ratings yet
- Plant Physiology IDocument7 pagesPlant Physiology IC'estMoiNo ratings yet
- Cell Wall: Presented by M. Vijaya LakshmiDocument9 pagesCell Wall: Presented by M. Vijaya LakshmiATCHUNALA SAINo ratings yet
- Cell PlantDocument7 pagesCell PlantEbiz MarkNo ratings yet
- Parts of The Cell WallDocument28 pagesParts of The Cell WallJOSHUA DICHOSONo ratings yet
- 3 Cytology Membran JunctionDocument48 pages3 Cytology Membran JunctionSamNo ratings yet
- Cover PageDocument7 pagesCover PageKamran AhmadNo ratings yet
- CellDocument16 pagesCellanantchouhdary1709No ratings yet
- 3 B Cytology-Membrane StructuresDocument19 pages3 B Cytology-Membrane Structuresm.mansoor3377No ratings yet
- Cell Walls: Structure, Biogenesis, and ExpansionDocument26 pagesCell Walls: Structure, Biogenesis, and ExpansionRavindraNo ratings yet
- Cell WallDocument5 pagesCell WallApril FrigillanaNo ratings yet
- Cell FunctionDocument26 pagesCell Functionreyesmarquis6uNo ratings yet
- Cellulose SynthesisDocument15 pagesCellulose SynthesisDanielNo ratings yet
- FT1043 Lecture 1Document40 pagesFT1043 Lecture 1Norzhafarina M. IsmailNo ratings yet
- Plant Cell WallDocument2 pagesPlant Cell WallShiela BelandresNo ratings yet
- Plant Structure and FunctionDocument165 pagesPlant Structure and Functionsalma.abumariamNo ratings yet
- Cell WallDocument29 pagesCell Wallkesiya franklinNo ratings yet
- Chapter 3 Micropara (Outline)Document5 pagesChapter 3 Micropara (Outline)Jezrylle BalaongNo ratings yet
- Presentation For Reporting in BiologyDocument61 pagesPresentation For Reporting in BiologyLovely Nhel EslomotNo ratings yet
- Cells Cell Membrane Over-Expansion Cell Plants Fungi Prokaryotic MycoplasmasDocument5 pagesCells Cell Membrane Over-Expansion Cell Plants Fungi Prokaryotic MycoplasmasbbhoeeNo ratings yet
- Cell MembraneDocument15 pagesCell MembraneAyesha SaleemNo ratings yet
- Cell Wall: by Fajrie AchlanaDocument35 pagesCell Wall: by Fajrie Achlanafajrieachlana100% (1)
- Parts and Function of Plant CellDocument3 pagesParts and Function of Plant CellAmy A. OgnayonNo ratings yet
- Plant Cytoskeleton PDFDocument7 pagesPlant Cytoskeleton PDFmanoj_rkl_07No ratings yet
- Study of Cellwall Constituents and Cell InclusionsDocument42 pagesStudy of Cellwall Constituents and Cell Inclusionsharshe v73% (11)
- Lecture 1 Cell and Molecular Biology 1Document38 pagesLecture 1 Cell and Molecular Biology 1Martin HangasNo ratings yet
- Plant Cell Structure & UltrastructureDocument73 pagesPlant Cell Structure & Ultrastructuresalma80877No ratings yet
- The Difference Between Animal and Plant Cell: ChloroplastsDocument6 pagesThe Difference Between Animal and Plant Cell: ChloroplastsYukiNo ratings yet
- Cell Theory: Functions of LifeDocument12 pagesCell Theory: Functions of LifeSeila MoriNo ratings yet
- Cellular Ultrastructure: Eukaryotic CellsDocument9 pagesCellular Ultrastructure: Eukaryotic CellsPiriyatharshini RamanathNo ratings yet
- Plant Cell OrganellesDocument13 pagesPlant Cell OrganellesKritika MalhotraNo ratings yet
- Cell Biology and Membrane BiochemistryDocument106 pagesCell Biology and Membrane BiochemistryVijay Bhasker TekulapallyNo ratings yet
- Plasmodesmata and VacuoleDocument27 pagesPlasmodesmata and VacuoleDeepak ThakurNo ratings yet
- Plant Cell WallDocument8 pagesPlant Cell WallErika MatiasNo ratings yet
- CELL Study MaterialDocument12 pagesCELL Study MaterialShyamasree SenguptaNo ratings yet
- Cell Wall SNDocument3 pagesCell Wall SNsamy gNo ratings yet
- Plant Cells and Plant Structure: AS Biology Unit 2 - Topic 4Document33 pagesPlant Cells and Plant Structure: AS Biology Unit 2 - Topic 4Escape VelocityNo ratings yet
- Cell Structure Function and PropertiesDocument10 pagesCell Structure Function and PropertiesWanda JohnNo ratings yet
- Pharmacognosy L02Document21 pagesPharmacognosy L02Youssef ElhamamyNo ratings yet
- Unit 3. Cellular Basis of Life-2Document13 pagesUnit 3. Cellular Basis of Life-2Kerala MekuriyaNo ratings yet
- Chapter 4 Nature and Composition of Crops 124136Document79 pagesChapter 4 Nature and Composition of Crops 124136S L A YNo ratings yet
- For Ntse CellDocument73 pagesFor Ntse CellPrakhar RajNo ratings yet
- Cell MembraneDocument7 pagesCell MembraneRishikesh BhintadeNo ratings yet
- Membrane and Cell Wall Deposition at The Division SiteDocument2 pagesMembrane and Cell Wall Deposition at The Division SiteNatalia LadinoNo ratings yet
- Lecture 1 - Cells Structure and Transport MechanismssdfkgjkDocument12 pagesLecture 1 - Cells Structure and Transport Mechanismssdfkgjktanique.nembhard1022No ratings yet
- The CellDocument20 pagesThe CellganeshchandraroutrayNo ratings yet
- Summative Test Reviewer C D E G K LDocument15 pagesSummative Test Reviewer C D E G K LGabriel John JueleNo ratings yet
- Cell Wall StructureDocument2 pagesCell Wall StructureDl Al-azizNo ratings yet
- 1st Year Chapter 1 Biology Book Fed BoardDocument86 pages1st Year Chapter 1 Biology Book Fed Boardwondersofworld.30No ratings yet
- Saccharomyces Cerevisiae YeastDocument11 pagesSaccharomyces Cerevisiae YeastNguyễn ThuNo ratings yet
- Presented By-Mohammad Zainul PG First YearDocument56 pagesPresented By-Mohammad Zainul PG First YearLAVKESH KUMARNo ratings yet
- Chemical Nature of The Cell WallDocument5 pagesChemical Nature of The Cell WallSanjoy NingthoujamNo ratings yet
- From Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.From EverandFrom Tubulin to Thought: The Nexus of Cytoskeleton Microtubules and Brain Complexity.No ratings yet
- Plant Cell Biology #2Document34 pagesPlant Cell Biology #2Anirudh Moza 16009No ratings yet
- Restoration Ecology PrimerDocument7 pagesRestoration Ecology PrimerAnirudh Moza 16009No ratings yet
- Magnetic Moment PDFDocument5 pagesMagnetic Moment PDFAnirudh Moza 16009No ratings yet
- Analysis & Diode Characterization of Mos Based Dna HydrogelDocument28 pagesAnalysis & Diode Characterization of Mos Based Dna HydrogelAnirudh Moza 16009No ratings yet
- Drain Cleaner Us 5783537a PDFDocument23 pagesDrain Cleaner Us 5783537a PDFLuis UlakiaNo ratings yet
- Economics and Environmental Impact of Bioethanol Production Technologies: An AppraisalDocument19 pagesEconomics and Environmental Impact of Bioethanol Production Technologies: An AppraisalChinwuba Samuel EbukaNo ratings yet
- Characterisation Studies and Impact of Chemical Treatment On Mechanical Properties of Sisal FiberDocument16 pagesCharacterisation Studies and Impact of Chemical Treatment On Mechanical Properties of Sisal FiberKebede kasaNo ratings yet
- Biochemical Engineering Fundamentals - Parte 2Document120 pagesBiochemical Engineering Fundamentals - Parte 2Jose Suarez100% (1)
- F5MR ch1 (1 28)Document28 pagesF5MR ch1 (1 28)NurulFaiza MohdNordinNo ratings yet
- Dietray Fibers (Complete Lecture)Document60 pagesDietray Fibers (Complete Lecture)Nauman SattarNo ratings yet
- Cotton FibreDocument5 pagesCotton FibreSieed HassanNo ratings yet
- Ebook Fibre Analysis of Animal Feed GB PDFDocument31 pagesEbook Fibre Analysis of Animal Feed GB PDFVlad MateiuNo ratings yet
- Characterization of Lignin Extracted CompoundDocument13 pagesCharacterization of Lignin Extracted CompoundJitender SinghNo ratings yet
- 2011 Chemistry NotesDocument70 pages2011 Chemistry NotesAbdullionNo ratings yet
- Exam 1 AnswersDocument9 pagesExam 1 AnswersA'Khris Fell-For Your-TypeNo ratings yet
- Echavia Concept-PaperDocument3 pagesEchavia Concept-PaperJefrey M. ButilNo ratings yet
- Cellulose PDFDocument10 pagesCellulose PDFclaudia estupiñanNo ratings yet
- Unit 4 GasesDocument6 pagesUnit 4 GasesLhenie DaquizNo ratings yet
- Sizing Tencel ModalDocument1 pageSizing Tencel ModalMahfuzNo ratings yet
- St. John The Baptist Catholic SchoolDocument3 pagesSt. John The Baptist Catholic SchoolJannelle ManarangNo ratings yet
- 10.1007@s40883 019 00134 1 PDFDocument24 pages10.1007@s40883 019 00134 1 PDFRohit GNo ratings yet
- Cellulose Fibers, RegeneratedDocument39 pagesCellulose Fibers, Regeneratedyemresimsek100% (1)
- Carbohydrates (Class 4)Document19 pagesCarbohydrates (Class 4)Swetank SahaiNo ratings yet
- Rangkuman Stock KainDocument1,142 pagesRangkuman Stock KainxtephenzNo ratings yet
- Carbohydrates For UGDocument139 pagesCarbohydrates For UGSantosh Kumar0% (1)
- Cambridge International AS and A Level Biology WorkbookDocument254 pagesCambridge International AS and A Level Biology WorkbookMash GrigNo ratings yet
- Heavy Metal Adsorption Onto Agro-Based Waste Materials - A ReviewDocument10 pagesHeavy Metal Adsorption Onto Agro-Based Waste Materials - A ReviewClaoston Nardon0% (1)
- Advances in Sustainable Polymers: Vimal Katiyar Amit Kumar Neha Mulchandani EditorsDocument421 pagesAdvances in Sustainable Polymers: Vimal Katiyar Amit Kumar Neha Mulchandani EditorsRahaf AljboriNo ratings yet
- 18cho101t Unit 3 Biomass EnergyDocument66 pages18cho101t Unit 3 Biomass EnergyHruday CoolkidNo ratings yet
- Cotton & JuteDocument5 pagesCotton & JuteSakshi SargarNo ratings yet
- Berkeley Science Review - Spring 2012Document56 pagesBerkeley Science Review - Spring 2012The Berkeley Science Review100% (1)
- Banana Pseudo Stem CHAPTER 1 3Document53 pagesBanana Pseudo Stem CHAPTER 1 3mizzpain CabuyadaoNo ratings yet
- Characterization of Refined Carrageenan From Kappaphycus AlvareziiDocument9 pagesCharacterization of Refined Carrageenan From Kappaphycus AlvareziiaviNo ratings yet