
Arabinose + Tryptophan Operon
Arabinose + Tryptophan Operon
You might also like
- 7Document15 pages7AmaniNo ratings yet
- WHO-EU Comparative Drinking Water StandardsDocument4 pagesWHO-EU Comparative Drinking Water StandardsLucy ZuluNo ratings yet
- Biochemistry - A Short CourseDocument1,026 pagesBiochemistry - A Short CourseFelicita Urzi100% (3)
- 07 Control of Gene Expression in ProkaryotesDocument17 pages07 Control of Gene Expression in Prokaryotesmustafa aliNo ratings yet
- Introduction To Genetic Analysis: Regulation of Gene Expression in Bacteria and Their VirusesDocument46 pagesIntroduction To Genetic Analysis: Regulation of Gene Expression in Bacteria and Their VirusesMuneeb ArshadNo ratings yet
- Chromosomal Map Begins at Oric Units of MinutesDocument10 pagesChromosomal Map Begins at Oric Units of Minutesdhita malikaNo ratings yet
- Arabinose Operon RI2022 Prelim P2Q4Document5 pagesArabinose Operon RI2022 Prelim P2Q4deekshana gouthamNo ratings yet
- 11 Biosynthesis of RNADocument52 pages11 Biosynthesis of RNADayne Ocampo-SolimanNo ratings yet
- Control of Gene Expression in BacteriaDocument18 pagesControl of Gene Expression in BacteriaFenamae EstorqueNo ratings yet
- Appendix AraOperonDocument2 pagesAppendix AraOperonhodaputhaNo ratings yet
- Base Alterations and Base DamageDocument22 pagesBase Alterations and Base Damagemasthan6yNo ratings yet
- 10 - Lac-OperonDocument20 pages10 - Lac-OperonAisha ranaNo ratings yet
- Gene RegulationDocument13 pagesGene RegulationLeenakshi TyagiNo ratings yet
- Regulation of Gene Expression-1Document30 pagesRegulation of Gene Expression-1mus zaharaNo ratings yet
- Biochem AssDocument9 pagesBiochem AssBereket AntenehNo ratings yet
- Wa0008.Document17 pagesWa0008.Rita JaypuriaNo ratings yet
- Gene Regulation in Prokaryotes-1Document17 pagesGene Regulation in Prokaryotes-1Fasiha Mushadi100% (1)
- 6 Lecture-TranslationDocument25 pages6 Lecture-Translationmoazzanrana678No ratings yet
- Prokaryote Gene Regulation 2023 2024Document33 pagesProkaryote Gene Regulation 2023 2024Thái AnNo ratings yet
- GENETICS Chapter 9 - From Gene To Protein - Genetic Code and Translation - EDITINGDocument53 pagesGENETICS Chapter 9 - From Gene To Protein - Genetic Code and Translation - EDITINGdo hieuNo ratings yet
- Regulation of Gene Expression in Prokaryotic and Eukaryotic CellsDocument13 pagesRegulation of Gene Expression in Prokaryotic and Eukaryotic CellsMifta KhuljannahNo ratings yet
- TranslationDocument44 pagesTranslationAnne Gabrielle Marapon AgsunodNo ratings yet
- Control of Gene Expression Part 1Document45 pagesControl of Gene Expression Part 1jenny joseNo ratings yet
- Lac Operon: O1 Is The Main Operator Site and Must Be Bound by The Repressor If Transcription IsDocument7 pagesLac Operon: O1 Is The Main Operator Site and Must Be Bound by The Repressor If Transcription IsShalmali ChatterjeeNo ratings yet
- The Operon ModelDocument44 pagesThe Operon Modelfae-ar_raziNo ratings yet
- Gene Expression, RNA, Transcription and Translation: Higher Human BiologyDocument58 pagesGene Expression, RNA, Transcription and Translation: Higher Human BiologyShruthiNo ratings yet
- Coli. Arabinose Must First Be Converted Into Ribulose-5-Phosphate Before It Can BeDocument5 pagesColi. Arabinose Must First Be Converted Into Ribulose-5-Phosphate Before It Can BeDiego LeonardoNo ratings yet
- Meredith Barb Homework-2 (25 Points) Due On Sept. 24 by 5:00pmDocument5 pagesMeredith Barb Homework-2 (25 Points) Due On Sept. 24 by 5:00pmMeredith BarbNo ratings yet
- Transcription Prokaryotes (RNAP, Promoter, Transcription InitiationDocument27 pagesTranscription Prokaryotes (RNAP, Promoter, Transcription Initiationnajla 2002No ratings yet
- RNA Synthesis and ProcessingDocument17 pagesRNA Synthesis and ProcessingInnocent Clifford MaranduNo ratings yet
- Eukaryote Translation PDFDocument21 pagesEukaryote Translation PDFChandra Mohan Meena100% (1)
- Lecture On Transcription and TranslationDocument47 pagesLecture On Transcription and TranslationAnna Beatrice BautistaNo ratings yet
- Gene RegulationDocument4 pagesGene RegulationPetros MaheyaNo ratings yet
- 5 Molecular Mechanisms of Gene RegulationDocument44 pages5 Molecular Mechanisms of Gene Regulationpramudi madushaniNo ratings yet
- TranslationDocument44 pagesTranslationlynzel Marie LozaresNo ratings yet
- GOF Unit3 Part 1Document24 pagesGOF Unit3 Part 1alshads957No ratings yet
- BCMB2x01/MEDS2003 Prokaryotic Transcription: Giselle - Yeo@sydney - Edu.auDocument25 pagesBCMB2x01/MEDS2003 Prokaryotic Transcription: Giselle - Yeo@sydney - Edu.auhyy370815No ratings yet
- The Arabinose Operon: Structure and MechanismDocument3 pagesThe Arabinose Operon: Structure and MechanismFildzah AdanyNo ratings yet
- Lect 4 Prokaryotic Gene StructureDocument45 pagesLect 4 Prokaryotic Gene StructureElias LibayNo ratings yet
- Gene Regulation - Lac Operon & TRP OperonDocument25 pagesGene Regulation - Lac Operon & TRP OperonSalam Pradeep Singh75% (12)
- 25 LN RNA Organization and Transcription CNRAADocument55 pages25 LN RNA Organization and Transcription CNRAADakshitha DharmakeerthiNo ratings yet
- Chapter 32 Lippincott BiochemistryDocument35 pagesChapter 32 Lippincott BiochemistryMeysam SajjadiNo ratings yet
- Molecular Regulation1Document23 pagesMolecular Regulation1Atharva PurohitNo ratings yet
- Gene Regulation: Mrs. Ofelia Solano SaludarDocument39 pagesGene Regulation: Mrs. Ofelia Solano SaludarmskikiNo ratings yet
- The Arabinose OperonDocument2 pagesThe Arabinose OperonSayan BhattacharyaNo ratings yet
- Transcription and RegulationDocument40 pagesTranscription and RegulationRUDRANSH PUJARINo ratings yet
- BP605T Unit 4-6Document482 pagesBP605T Unit 4-6Gyampoh SolomonNo ratings yet
- Complex Regulation of The Ara OperonDocument12 pagesComplex Regulation of The Ara OperonFemi Mega LestariNo ratings yet
- Lecture 2Document33 pagesLecture 2SUNDAS FATIMANo ratings yet
- Gene ExpressionDocument72 pagesGene ExpressionMika ForwardNo ratings yet
- Gene Regulation PDFDocument49 pagesGene Regulation PDFGabriel NdandaniNo ratings yet
- Genetik MikrobDocument33 pagesGenetik MikrobekasukmawatyNo ratings yet
- 106 Ceng w10Document62 pages106 Ceng w10mertNo ratings yet
- Transciption and TranslationDocument31 pagesTransciption and TranslationDuduetsang MosalakataneNo ratings yet
- Regulation of Lactose Metabolism (Positive and Negative) : Molecular Biology AssignmentDocument20 pagesRegulation of Lactose Metabolism (Positive and Negative) : Molecular Biology AssignmentswarNo ratings yet
- Regulation of Gene Expression in Prokaryotes (Lac Operon)Document13 pagesRegulation of Gene Expression in Prokaryotes (Lac Operon)MUHAMMAD ANEEQ JAVEDNo ratings yet
- Protein SynthesisDocument54 pagesProtein SynthesisGemmie Jn PierreNo ratings yet
- Translation 160204065032Document79 pagesTranslation 160204065032Sai SundeepNo ratings yet
- Transcription Prokaryotes 2012-cDocument27 pagesTranscription Prokaryotes 2012-cAnupama PatiNo ratings yet
- Lecture8 DNA-Dependent RNA Synthesis-2Document50 pagesLecture8 DNA-Dependent RNA Synthesis-2Constance WongNo ratings yet
- In ClassWorksheetDocument4 pagesIn ClassWorksheetSZNo ratings yet
- RegGenExp (Khusus Palangkaraya 2011), Bahan Ujian SajaDocument20 pagesRegGenExp (Khusus Palangkaraya 2011), Bahan Ujian SajaeciNo ratings yet
- Isolation and Spectrophotometric Characterization o F Photosynthetic Pigments Rodney F BoyerDocument4 pagesIsolation and Spectrophotometric Characterization o F Photosynthetic Pigments Rodney F BoyerLucy ZuluNo ratings yet
- Lecture:6 Food Microbiology Microbial Spoilage of Cereals and Bakery FoodsDocument4 pagesLecture:6 Food Microbiology Microbial Spoilage of Cereals and Bakery FoodsLucy ZuluNo ratings yet
- Identification of Microorganisms Responsible For S PDFDocument3 pagesIdentification of Microorganisms Responsible For S PDFLucy ZuluNo ratings yet
- Che2522 Test 2Document3 pagesChe2522 Test 2Lucy ZuluNo ratings yet
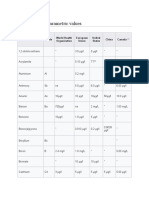
- Comparison of Parametric ValuesDocument5 pagesComparison of Parametric ValuesLucy ZuluNo ratings yet
- Cheese Production BIO 4352: DR Sydney MalamaDocument3 pagesCheese Production BIO 4352: DR Sydney MalamaLucy ZuluNo ratings yet
- BIO 4352 - Ass - 1 - 2020 PDFDocument1 pageBIO 4352 - Ass - 1 - 2020 PDFLucy ZuluNo ratings yet
- Carbohydrate MetabolismDocument52 pagesCarbohydrate MetabolismLucy ZuluNo ratings yet
- PDF PDFDocument31 pagesPDF PDFLucy ZuluNo ratings yet
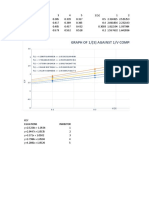
- Graph of 1/ (S) Against 1/V Competive InhibitionDocument2 pagesGraph of 1/ (S) Against 1/V Competive InhibitionLucy ZuluNo ratings yet
- Translation Initiation: April 2017 BIO 3421Document109 pagesTranslation Initiation: April 2017 BIO 3421Lucy ZuluNo ratings yet
- CHE 3122 Lecture Notes Energetics of Photosynthesis: 2018 Academic Year October 2019Document9 pagesCHE 3122 Lecture Notes Energetics of Photosynthesis: 2018 Academic Year October 2019Lucy ZuluNo ratings yet
- Che 3111 Lecture Notes, Nitrogen Metabolism 2019 July 2020 PDFDocument34 pagesChe 3111 Lecture Notes, Nitrogen Metabolism 2019 July 2020 PDFLucy ZuluNo ratings yet
- CHE2522 Course Outline - 2019-2020 Academic Year-Final 24.07.2020Document5 pagesCHE2522 Course Outline - 2019-2020 Academic Year-Final 24.07.2020Lucy ZuluNo ratings yet
- Graph of 1/ (S) Against 1/V Competive InhibitionDocument2 pagesGraph of 1/ (S) Against 1/V Competive InhibitionLucy ZuluNo ratings yet
- The University of Zambia School of Natural Sciences Department of Biological Sciences BIO-4321Document4 pagesThe University of Zambia School of Natural Sciences Department of Biological Sciences BIO-4321Lucy ZuluNo ratings yet
- Lipid Metabolism NotesDocument17 pagesLipid Metabolism NotesLucy ZuluNo ratings yet
- Coli. Arabinose Must First Be Converted Into Ribulose-5-Phosphate Before It Can BeDocument5 pagesColi. Arabinose Must First Be Converted Into Ribulose-5-Phosphate Before It Can BeDiego LeonardoNo ratings yet
- Student Manual pGLO Transformation: Lesson 1 Introduction To TransformationDocument26 pagesStudent Manual pGLO Transformation: Lesson 1 Introduction To TransformationanamqadirNo ratings yet
- Bacillus Megaterium: Protein Production SystemDocument35 pagesBacillus Megaterium: Protein Production SystemNurul Apsari AjiNo ratings yet
- Fundamental Molecular Biology: Lisabeth A. AllisonDocument96 pagesFundamental Molecular Biology: Lisabeth A. AllisonJu noNo ratings yet
- 2 Regulation Gene Expression BalaccoDocument34 pages2 Regulation Gene Expression Balacco9147Indah WulandariNo ratings yet
- SpGLOTransformation PDFDocument16 pagesSpGLOTransformation PDFaspergillus_jallNo ratings yet
- The Arabinose OperonDocument2 pagesThe Arabinose OperonSayan BhattacharyaNo ratings yet
- Ara Operon Notes - 18Document4 pagesAra Operon Notes - 18Khaleeda Ahmad AnuarNo ratings yet
- BLG143 - Final Exam Cheat SheetDocument2 pagesBLG143 - Final Exam Cheat SheetIndraragini AnpalakanNo ratings yet
- Arabinose + Tryptophan OperonDocument58 pagesArabinose + Tryptophan OperonLucy ZuluNo ratings yet






















































































Download as pptx, pdf, or txt
You might also like
- 7Document15 pages7AmaniNo ratings yet
- WHO-EU Comparative Drinking Water StandardsDocument4 pagesWHO-EU Comparative Drinking Water StandardsLucy ZuluNo ratings yet
- Biochemistry - A Short CourseDocument1,026 pagesBiochemistry - A Short CourseFelicita Urzi100% (3)
- 07 Control of Gene Expression in ProkaryotesDocument17 pages07 Control of Gene Expression in Prokaryotesmustafa aliNo ratings yet
- Introduction To Genetic Analysis: Regulation of Gene Expression in Bacteria and Their VirusesDocument46 pagesIntroduction To Genetic Analysis: Regulation of Gene Expression in Bacteria and Their VirusesMuneeb ArshadNo ratings yet
- Chromosomal Map Begins at Oric Units of MinutesDocument10 pagesChromosomal Map Begins at Oric Units of Minutesdhita malikaNo ratings yet
- Arabinose Operon RI2022 Prelim P2Q4Document5 pagesArabinose Operon RI2022 Prelim P2Q4deekshana gouthamNo ratings yet
- 11 Biosynthesis of RNADocument52 pages11 Biosynthesis of RNADayne Ocampo-SolimanNo ratings yet
- Control of Gene Expression in BacteriaDocument18 pagesControl of Gene Expression in BacteriaFenamae EstorqueNo ratings yet
- Appendix AraOperonDocument2 pagesAppendix AraOperonhodaputhaNo ratings yet
- Base Alterations and Base DamageDocument22 pagesBase Alterations and Base Damagemasthan6yNo ratings yet
- 10 - Lac-OperonDocument20 pages10 - Lac-OperonAisha ranaNo ratings yet
- Gene RegulationDocument13 pagesGene RegulationLeenakshi TyagiNo ratings yet
- Regulation of Gene Expression-1Document30 pagesRegulation of Gene Expression-1mus zaharaNo ratings yet
- Biochem AssDocument9 pagesBiochem AssBereket AntenehNo ratings yet
- Wa0008.Document17 pagesWa0008.Rita JaypuriaNo ratings yet
- Gene Regulation in Prokaryotes-1Document17 pagesGene Regulation in Prokaryotes-1Fasiha Mushadi100% (1)
- 6 Lecture-TranslationDocument25 pages6 Lecture-Translationmoazzanrana678No ratings yet
- Prokaryote Gene Regulation 2023 2024Document33 pagesProkaryote Gene Regulation 2023 2024Thái AnNo ratings yet
- GENETICS Chapter 9 - From Gene To Protein - Genetic Code and Translation - EDITINGDocument53 pagesGENETICS Chapter 9 - From Gene To Protein - Genetic Code and Translation - EDITINGdo hieuNo ratings yet
- Regulation of Gene Expression in Prokaryotic and Eukaryotic CellsDocument13 pagesRegulation of Gene Expression in Prokaryotic and Eukaryotic CellsMifta KhuljannahNo ratings yet
- TranslationDocument44 pagesTranslationAnne Gabrielle Marapon AgsunodNo ratings yet
- Control of Gene Expression Part 1Document45 pagesControl of Gene Expression Part 1jenny joseNo ratings yet
- Lac Operon: O1 Is The Main Operator Site and Must Be Bound by The Repressor If Transcription IsDocument7 pagesLac Operon: O1 Is The Main Operator Site and Must Be Bound by The Repressor If Transcription IsShalmali ChatterjeeNo ratings yet
- The Operon ModelDocument44 pagesThe Operon Modelfae-ar_raziNo ratings yet
- Gene Expression, RNA, Transcription and Translation: Higher Human BiologyDocument58 pagesGene Expression, RNA, Transcription and Translation: Higher Human BiologyShruthiNo ratings yet
- Coli. Arabinose Must First Be Converted Into Ribulose-5-Phosphate Before It Can BeDocument5 pagesColi. Arabinose Must First Be Converted Into Ribulose-5-Phosphate Before It Can BeDiego LeonardoNo ratings yet
- Meredith Barb Homework-2 (25 Points) Due On Sept. 24 by 5:00pmDocument5 pagesMeredith Barb Homework-2 (25 Points) Due On Sept. 24 by 5:00pmMeredith BarbNo ratings yet
- Transcription Prokaryotes (RNAP, Promoter, Transcription InitiationDocument27 pagesTranscription Prokaryotes (RNAP, Promoter, Transcription Initiationnajla 2002No ratings yet
- RNA Synthesis and ProcessingDocument17 pagesRNA Synthesis and ProcessingInnocent Clifford MaranduNo ratings yet
- Eukaryote Translation PDFDocument21 pagesEukaryote Translation PDFChandra Mohan Meena100% (1)
- Lecture On Transcription and TranslationDocument47 pagesLecture On Transcription and TranslationAnna Beatrice BautistaNo ratings yet
- Gene RegulationDocument4 pagesGene RegulationPetros MaheyaNo ratings yet
- 5 Molecular Mechanisms of Gene RegulationDocument44 pages5 Molecular Mechanisms of Gene Regulationpramudi madushaniNo ratings yet
- TranslationDocument44 pagesTranslationlynzel Marie LozaresNo ratings yet
- GOF Unit3 Part 1Document24 pagesGOF Unit3 Part 1alshads957No ratings yet
- BCMB2x01/MEDS2003 Prokaryotic Transcription: Giselle - Yeo@sydney - Edu.auDocument25 pagesBCMB2x01/MEDS2003 Prokaryotic Transcription: Giselle - Yeo@sydney - Edu.auhyy370815No ratings yet
- The Arabinose Operon: Structure and MechanismDocument3 pagesThe Arabinose Operon: Structure and MechanismFildzah AdanyNo ratings yet
- Lect 4 Prokaryotic Gene StructureDocument45 pagesLect 4 Prokaryotic Gene StructureElias LibayNo ratings yet
- Gene Regulation - Lac Operon & TRP OperonDocument25 pagesGene Regulation - Lac Operon & TRP OperonSalam Pradeep Singh75% (12)
- 25 LN RNA Organization and Transcription CNRAADocument55 pages25 LN RNA Organization and Transcription CNRAADakshitha DharmakeerthiNo ratings yet
- Chapter 32 Lippincott BiochemistryDocument35 pagesChapter 32 Lippincott BiochemistryMeysam SajjadiNo ratings yet
- Molecular Regulation1Document23 pagesMolecular Regulation1Atharva PurohitNo ratings yet
- Gene Regulation: Mrs. Ofelia Solano SaludarDocument39 pagesGene Regulation: Mrs. Ofelia Solano SaludarmskikiNo ratings yet
- The Arabinose OperonDocument2 pagesThe Arabinose OperonSayan BhattacharyaNo ratings yet
- Transcription and RegulationDocument40 pagesTranscription and RegulationRUDRANSH PUJARINo ratings yet
- BP605T Unit 4-6Document482 pagesBP605T Unit 4-6Gyampoh SolomonNo ratings yet
- Complex Regulation of The Ara OperonDocument12 pagesComplex Regulation of The Ara OperonFemi Mega LestariNo ratings yet
- Lecture 2Document33 pagesLecture 2SUNDAS FATIMANo ratings yet
- Gene ExpressionDocument72 pagesGene ExpressionMika ForwardNo ratings yet
- Gene Regulation PDFDocument49 pagesGene Regulation PDFGabriel NdandaniNo ratings yet
- Genetik MikrobDocument33 pagesGenetik MikrobekasukmawatyNo ratings yet
- 106 Ceng w10Document62 pages106 Ceng w10mertNo ratings yet
- Transciption and TranslationDocument31 pagesTransciption and TranslationDuduetsang MosalakataneNo ratings yet
- Regulation of Lactose Metabolism (Positive and Negative) : Molecular Biology AssignmentDocument20 pagesRegulation of Lactose Metabolism (Positive and Negative) : Molecular Biology AssignmentswarNo ratings yet
- Regulation of Gene Expression in Prokaryotes (Lac Operon)Document13 pagesRegulation of Gene Expression in Prokaryotes (Lac Operon)MUHAMMAD ANEEQ JAVEDNo ratings yet
- Protein SynthesisDocument54 pagesProtein SynthesisGemmie Jn PierreNo ratings yet
- Translation 160204065032Document79 pagesTranslation 160204065032Sai SundeepNo ratings yet
- Transcription Prokaryotes 2012-cDocument27 pagesTranscription Prokaryotes 2012-cAnupama PatiNo ratings yet
- Lecture8 DNA-Dependent RNA Synthesis-2Document50 pagesLecture8 DNA-Dependent RNA Synthesis-2Constance WongNo ratings yet
- In ClassWorksheetDocument4 pagesIn ClassWorksheetSZNo ratings yet
- RegGenExp (Khusus Palangkaraya 2011), Bahan Ujian SajaDocument20 pagesRegGenExp (Khusus Palangkaraya 2011), Bahan Ujian SajaeciNo ratings yet
- Isolation and Spectrophotometric Characterization o F Photosynthetic Pigments Rodney F BoyerDocument4 pagesIsolation and Spectrophotometric Characterization o F Photosynthetic Pigments Rodney F BoyerLucy ZuluNo ratings yet
- Lecture:6 Food Microbiology Microbial Spoilage of Cereals and Bakery FoodsDocument4 pagesLecture:6 Food Microbiology Microbial Spoilage of Cereals and Bakery FoodsLucy ZuluNo ratings yet
- Identification of Microorganisms Responsible For S PDFDocument3 pagesIdentification of Microorganisms Responsible For S PDFLucy ZuluNo ratings yet
- Che2522 Test 2Document3 pagesChe2522 Test 2Lucy ZuluNo ratings yet
- Comparison of Parametric ValuesDocument5 pagesComparison of Parametric ValuesLucy ZuluNo ratings yet
- Cheese Production BIO 4352: DR Sydney MalamaDocument3 pagesCheese Production BIO 4352: DR Sydney MalamaLucy ZuluNo ratings yet
- BIO 4352 - Ass - 1 - 2020 PDFDocument1 pageBIO 4352 - Ass - 1 - 2020 PDFLucy ZuluNo ratings yet
- Carbohydrate MetabolismDocument52 pagesCarbohydrate MetabolismLucy ZuluNo ratings yet
- PDF PDFDocument31 pagesPDF PDFLucy ZuluNo ratings yet
- Graph of 1/ (S) Against 1/V Competive InhibitionDocument2 pagesGraph of 1/ (S) Against 1/V Competive InhibitionLucy ZuluNo ratings yet
- Translation Initiation: April 2017 BIO 3421Document109 pagesTranslation Initiation: April 2017 BIO 3421Lucy ZuluNo ratings yet
- CHE 3122 Lecture Notes Energetics of Photosynthesis: 2018 Academic Year October 2019Document9 pagesCHE 3122 Lecture Notes Energetics of Photosynthesis: 2018 Academic Year October 2019Lucy ZuluNo ratings yet
- Che 3111 Lecture Notes, Nitrogen Metabolism 2019 July 2020 PDFDocument34 pagesChe 3111 Lecture Notes, Nitrogen Metabolism 2019 July 2020 PDFLucy ZuluNo ratings yet
- CHE2522 Course Outline - 2019-2020 Academic Year-Final 24.07.2020Document5 pagesCHE2522 Course Outline - 2019-2020 Academic Year-Final 24.07.2020Lucy ZuluNo ratings yet
- Graph of 1/ (S) Against 1/V Competive InhibitionDocument2 pagesGraph of 1/ (S) Against 1/V Competive InhibitionLucy ZuluNo ratings yet
- The University of Zambia School of Natural Sciences Department of Biological Sciences BIO-4321Document4 pagesThe University of Zambia School of Natural Sciences Department of Biological Sciences BIO-4321Lucy ZuluNo ratings yet
- Lipid Metabolism NotesDocument17 pagesLipid Metabolism NotesLucy ZuluNo ratings yet
- Coli. Arabinose Must First Be Converted Into Ribulose-5-Phosphate Before It Can BeDocument5 pagesColi. Arabinose Must First Be Converted Into Ribulose-5-Phosphate Before It Can BeDiego LeonardoNo ratings yet
- Student Manual pGLO Transformation: Lesson 1 Introduction To TransformationDocument26 pagesStudent Manual pGLO Transformation: Lesson 1 Introduction To TransformationanamqadirNo ratings yet
- Bacillus Megaterium: Protein Production SystemDocument35 pagesBacillus Megaterium: Protein Production SystemNurul Apsari AjiNo ratings yet
- Fundamental Molecular Biology: Lisabeth A. AllisonDocument96 pagesFundamental Molecular Biology: Lisabeth A. AllisonJu noNo ratings yet
- 2 Regulation Gene Expression BalaccoDocument34 pages2 Regulation Gene Expression Balacco9147Indah WulandariNo ratings yet
- SpGLOTransformation PDFDocument16 pagesSpGLOTransformation PDFaspergillus_jallNo ratings yet
- The Arabinose OperonDocument2 pagesThe Arabinose OperonSayan BhattacharyaNo ratings yet
- Ara Operon Notes - 18Document4 pagesAra Operon Notes - 18Khaleeda Ahmad AnuarNo ratings yet
- BLG143 - Final Exam Cheat SheetDocument2 pagesBLG143 - Final Exam Cheat SheetIndraragini AnpalakanNo ratings yet
- Arabinose + Tryptophan OperonDocument58 pagesArabinose + Tryptophan OperonLucy ZuluNo ratings yet