Download as pptx, pdf, or txt
You might also like
- NickBare S Cookbook 1.0Document35 pagesNickBare S Cookbook 1.0Panos VNo ratings yet
- Beta Oxidation of FattyacidDocument26 pagesBeta Oxidation of FattyacidLipsa Rani Panda100% (1)
- GlycogenolysisDocument37 pagesGlycogenolysisJasmine Rey QuintoNo ratings yet
- Chemistry of LipidsDocument123 pagesChemistry of LipidsLyra Get100% (1)
- Butter Cake (The Best!) PDFDocument3 pagesButter Cake (The Best!) PDFOblivionOmbre100% (1)
- Fatty Acid Synthesis 11.12.19Document18 pagesFatty Acid Synthesis 11.12.19Sanreet Randhawa100% (1)
- Chapter 21 Lipid BiosynthesisDocument44 pagesChapter 21 Lipid Biosynthesisapi-19916399No ratings yet
- Cholesterol - Synthesis, Metabolism, Regulation PDFDocument10 pagesCholesterol - Synthesis, Metabolism, Regulation PDFAdreiTheTripleA100% (1)
- Beta OxidationDocument19 pagesBeta Oxidationindra100% (1)
- Metabolism of Purine & Pyrimidine NucleotidesDocument38 pagesMetabolism of Purine & Pyrimidine NucleotidesShimmering MoonNo ratings yet
- 2 - Lipid Metabolism Lecture For StudentsDocument68 pages2 - Lipid Metabolism Lecture For StudentshwhsgxNo ratings yet
- Beta OxidationDocument25 pagesBeta OxidationManoj SigdelNo ratings yet
- Glycogenesis & GlycogenolysisDocument68 pagesGlycogenesis & GlycogenolysisAnn Michelle TarrobagoNo ratings yet
- HW - Carbohydrate Metabolism II & Lipid MetabolismDocument2 pagesHW - Carbohydrate Metabolism II & Lipid MetabolismyanNo ratings yet
- Lipid Profile L - DeterminatinDocument33 pagesLipid Profile L - Determinatinali100% (1)
- Metabolism of Ketone BodiesDocument38 pagesMetabolism of Ketone BodiesJavedNo ratings yet
- Fed State of MetabolismDocument40 pagesFed State of MetabolismBHARANIDHARAN M.VNo ratings yet
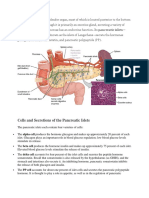
- Cells and Secretions of The Pancreatic IsletsDocument4 pagesCells and Secretions of The Pancreatic IsletsSophia OcayNo ratings yet
- CH 7 Notes Cellular RespirationDocument3 pagesCH 7 Notes Cellular RespirationCJ100% (1)
- Glucose, Part1Document33 pagesGlucose, Part1SarahNo ratings yet
- Also Known As TCA Cycle Essentially Involves This Is The Final Common Oxidative Pathway ForDocument10 pagesAlso Known As TCA Cycle Essentially Involves This Is The Final Common Oxidative Pathway ForNarayan debnath hridoyNo ratings yet
- Metabolism of LipidsDocument47 pagesMetabolism of LipidsMarwahNo ratings yet
- HMPDocument44 pagesHMPraanja2No ratings yet
- Metabolism and NutritionDocument52 pagesMetabolism and NutritionAlex SaljayNo ratings yet
- Hexose Monophosphate ShuntDocument18 pagesHexose Monophosphate ShuntAbdul Jabbar Abdul JabbarNo ratings yet
- Carbohydrates MetabolismDocument6 pagesCarbohydrates MetabolismAisha ShahfiqueeNo ratings yet
- HMP-shunt MEDDocument37 pagesHMP-shunt MEDAboubakar Moalim Mahad moh'dNo ratings yet
- Ketone Bodies 2012Document15 pagesKetone Bodies 2012Adhya MurugesanNo ratings yet
- Krebs CycleDocument2 pagesKrebs Cycleapi-327731714100% (1)
- Plasma ProteinDocument79 pagesPlasma ProteinnadieNo ratings yet
- Energy MetabolismDocument40 pagesEnergy MetabolismRuby JayaseelanNo ratings yet
- Digestive System NotesDocument10 pagesDigestive System NotesArchanna VyassNo ratings yet
- Lipoprotein Metabolism: PSC 3110 Fall 2004Document99 pagesLipoprotein Metabolism: PSC 3110 Fall 2004Irma MarianyNo ratings yet
- PROTEIN METABOLISM Dea Farha Fira Darson FineDocument44 pagesPROTEIN METABOLISM Dea Farha Fira Darson FineFarhati Mardhiyah100% (1)
- Metabolism of CarbohydrateDocument48 pagesMetabolism of CarbohydrateAbdullah TheNo ratings yet
- Hexose Mono Phosphate (HMP) ShuntDocument28 pagesHexose Mono Phosphate (HMP) ShuntVishesh JainNo ratings yet
- LipidsDocument13 pagesLipidsalianaNo ratings yet
- Ketone BodiesDocument16 pagesKetone BodiesAsim Ali100% (1)
- 1.carbohydrates and Lipid Metabolism-Converted - WatermarkDocument97 pages1.carbohydrates and Lipid Metabolism-Converted - WatermarkJuliyamol JoseNo ratings yet
- Gluconeogenesis, Glycogen MetabolismDocument35 pagesGluconeogenesis, Glycogen MetabolismdanielachynaNo ratings yet
- MCQ MembranesDocument10 pagesMCQ MembranesMarilyne RizkNo ratings yet
- Integration of Metabolism 2019Document68 pagesIntegration of Metabolism 2019Cahyani Tiara Safitri100% (1)
- Krebs Cycle GuideDocument3 pagesKrebs Cycle GuideMarie Sam100% (1)
- ProteinDocument13 pagesProteinapi-272775120No ratings yet
- Absorption of LipidsDocument22 pagesAbsorption of LipidsEmm Noman100% (1)
- Glomerular Function TestsDocument26 pagesGlomerular Function TestsAvi Verma100% (1)
- Nuclei C Acids: BSN 1y0-5Document25 pagesNuclei C Acids: BSN 1y0-5joevette_30No ratings yet
- GluconeogenesisDocument48 pagesGluconeogenesisQshyanNo ratings yet
- Carbohydrate, Lipid, Protein MetabolismDocument3 pagesCarbohydrate, Lipid, Protein Metabolismtritone.paradoxNo ratings yet
- Chemistryofproteinswithclinicalapplications 190621192525 PDFDocument181 pagesChemistryofproteinswithclinicalapplications 190621192525 PDFAl-waleed Julkanain100% (1)
- Lipid Profile: by Micheal and ChristianDocument15 pagesLipid Profile: by Micheal and ChristianMichealNo ratings yet
- Biochemistry II Ch. 16 Glycolysis and Gluconeogenesis (MASTER) )Document75 pagesBiochemistry II Ch. 16 Glycolysis and Gluconeogenesis (MASTER) )coco100% (1)
- Metabolism of Lipids: The Biochemistry and Molecular Biology Department of CMUDocument270 pagesMetabolism of Lipids: The Biochemistry and Molecular Biology Department of CMUanon_49132843No ratings yet
- Purine & Pyrimidine MetabolismDocument22 pagesPurine & Pyrimidine MetabolismManda100% (1)
- G6PD, GalactosemiaDocument32 pagesG6PD, GalactosemiaEricka B. BanaszczukNo ratings yet
- 2.metabolism of LipidsDocument159 pages2.metabolism of LipidsChristian Tham100% (1)
- GluconeogenesisDocument11 pagesGluconeogenesisMithilesh RautNo ratings yet
- CarbohydratemetabolismDocument88 pagesCarbohydratemetabolismEthio funNo ratings yet
- MCQ Carbohydrates Pdffor More JoinDocument6 pagesMCQ Carbohydrates Pdffor More JoinVikash KushwahaNo ratings yet
- Nucleic Acid DisordersDocument29 pagesNucleic Acid Disorderskritimahajan1989No ratings yet
- Carbohydrates MetabolismDocument152 pagesCarbohydrates Metabolismrio1995No ratings yet
- CHAPTER 24 Lipid BiosynthesisDocument16 pagesCHAPTER 24 Lipid Biosynthesis楊畯凱No ratings yet
- Fatty Acid SynthesisDocument28 pagesFatty Acid Synthesishassanainshahi13No ratings yet
- Metabolism of LipidsDocument98 pagesMetabolism of LipidsLyra GetNo ratings yet
- Lipid: Chemistry and MetabolismDocument68 pagesLipid: Chemistry and MetabolismLyra GetNo ratings yet
- Metabolism of LipidsDocument96 pagesMetabolism of LipidsLyra GetNo ratings yet
- 1 - 1 - Course OverviewDocument3 pages1 - 1 - Course OverviewchapincolNo ratings yet
- Yusuf Duale Update C.V - 11Document3 pagesYusuf Duale Update C.V - 11Yousuf DualeNo ratings yet
- Exporting Mango From Kenya To EuropeDocument57 pagesExporting Mango From Kenya To EuropeMohamedNo ratings yet
- Keto Meal Prep For US 2021 - A 10-Week Keto Meal Plan Guide With Recipes For Keto SuccessDocument151 pagesKeto Meal Prep For US 2021 - A 10-Week Keto Meal Plan Guide With Recipes For Keto Successmarc MacaaroNo ratings yet
- Indian Pregnancy Diet Plan For Women - Max HospitalDocument5 pagesIndian Pregnancy Diet Plan For Women - Max HospitalAzhar MandhraNo ratings yet
- Preparation of Moringa Capsules: Acta Scientific Agriculture (Issn: 2581-365X)Document3 pagesPreparation of Moringa Capsules: Acta Scientific Agriculture (Issn: 2581-365X)smail bendrissouNo ratings yet
- 10 Surefire Signs A Guy Is Flirting With YouDocument23 pages10 Surefire Signs A Guy Is Flirting With YouanaidaceNo ratings yet
- Oxalate-Rich FoodsDocument5 pagesOxalate-Rich FoodsRonNo ratings yet
- Wilson DiseaseDocument27 pagesWilson DiseaseDilshad TasnimNo ratings yet
- Meet The Meat Alternatives: The Value of Alternative Protein SourcesDocument10 pagesMeet The Meat Alternatives: The Value of Alternative Protein Sourcessiwei qinNo ratings yet
- FISH-BAR (Miyio)Document11 pagesFISH-BAR (Miyio)Ahmad DanishNo ratings yet
- Cupcake Bouquet Marketing PlanDocument9 pagesCupcake Bouquet Marketing PlanAel Scholar1No ratings yet
- Diet and Nutrition in Geriatric PatientsDocument38 pagesDiet and Nutrition in Geriatric PatientsDapinder BrarNo ratings yet
- Alternative Processing Machine For Swine Feed ProductionDocument10 pagesAlternative Processing Machine For Swine Feed Productionagility dreamsNo ratings yet
- Sugar Report WHO 107773 Updated and Revised Dec 2017Document65 pagesSugar Report WHO 107773 Updated and Revised Dec 2017Leonardo Moreno - ChacónNo ratings yet
- Biochemistry Midterm Carbohydrates: ClassificationsDocument21 pagesBiochemistry Midterm Carbohydrates: ClassificationsArah Lyn ApiagNo ratings yet
- Cookie BitesDocument3 pagesCookie BitesRicajean TaguranNo ratings yet
- Exploring Feeding BehaviorDocument6 pagesExploring Feeding BehaviorristeacristiNo ratings yet
- Best Macros Calculator For Tracking Muscle Gain and Fat Loss BarBendDocument1 pageBest Macros Calculator For Tracking Muscle Gain and Fat Loss BarBendTaimurMirzaNo ratings yet
- Uts Bahasa Inggris Ii Semester IiDocument4 pagesUts Bahasa Inggris Ii Semester Iibintang pamungkasNo ratings yet
- Pip Cordymax enDocument2 pagesPip Cordymax enIsa Lluïsa Palou EliasNo ratings yet
- Carb Cycling That Actually Works - BiotestDocument8 pagesCarb Cycling That Actually Works - BiotestDr. Abhijit Mande100% (2)
- Engineering Mechanics Dynamics 4Th Edition Pytel Solutions Manual Full Chapter PDFDocument27 pagesEngineering Mechanics Dynamics 4Th Edition Pytel Solutions Manual Full Chapter PDFrichard.eis609100% (12)
- PE Quiz CompilationDocument3 pagesPE Quiz CompilationSHEEN ALUBANo ratings yet
- Trees With Edible LeavesDocument80 pagesTrees With Edible LeavesGraham WattNo ratings yet
- CAPE Biology 2004 U2 P1Document20 pagesCAPE Biology 2004 U2 P1Sheniqua Greaves100% (1)
- Asignatura 6,0Document6 pagesAsignatura 6,0Dari Lenny Tavares ValdezNo ratings yet
- Cosmetics: Spirulina For Skin Care: A Bright Blue FutureDocument19 pagesCosmetics: Spirulina For Skin Care: A Bright Blue FutureEGA HPNo ratings yet