
Muscle Tissues: Mrs. Ofelia Solano Saludar
Muscle Tissues: Mrs. Ofelia Solano Saludar
You might also like
- Biomolecules Recap Answer Key by The Amoeba Sisters AnswerkeyDocument3 pagesBiomolecules Recap Answer Key by The Amoeba Sisters AnswerkeySandra Lopez50% (2)
- Muscular TissueDocument65 pagesMuscular TissueMitzel SapaloNo ratings yet
- Histology of MuscleDocument49 pagesHistology of MuscleTewachew Dessie100% (1)
- Muscle Tissue-Wl06.11Document54 pagesMuscle Tissue-Wl06.11api-19641337No ratings yet
- 29.-31. SBRC MSK SummaryDocument153 pages29.-31. SBRC MSK SummaryErin HillNo ratings yet
- Chap-20 - Locomotion & MovementDocument52 pagesChap-20 - Locomotion & MovementMittal SavaniNo ratings yet
- Muscular TDocument18 pagesMuscular TSumayya KabeerNo ratings yet
- MascularDocument39 pagesMascularAive BelistaNo ratings yet
- Muscle Tissue 2012Document13 pagesMuscle Tissue 2012ST. Fatimah KadirNo ratings yet
- FF Raumenys PDFDocument34 pagesFF Raumenys PDFLaura PaškevičiūtėNo ratings yet
- Muscle Tissue Lesson 1Document5 pagesMuscle Tissue Lesson 1Steves LeeboyNo ratings yet
- Muscle Tissue: - Skeletal Muscle - Cardiac Muscle - Smooth MuscleDocument36 pagesMuscle Tissue: - Skeletal Muscle - Cardiac Muscle - Smooth MuscleDentist Dina Samy100% (1)
- Muscular TissuesDocument46 pagesMuscular TissuesDAVE CANALETANo ratings yet
- Sweeney 2018Document14 pagesSweeney 2018LuisJauweriNo ratings yet
- 6.muscular TissueDocument40 pages6.muscular Tissuemero wowNo ratings yet
- 04muscle TissueDocument39 pages04muscle Tissueapi-19916399No ratings yet
- Histology of Cardiac and Smooth MuscleDocument73 pagesHistology of Cardiac and Smooth MuscleShehryar AbbasNo ratings yet
- Histology of Cardiac and Smooth MuscleDocument73 pagesHistology of Cardiac and Smooth MuscleShehryar AbbasNo ratings yet
- LPT 321Document103 pagesLPT 321rahul roshanNo ratings yet
- ANPH Midterm NotesDocument44 pagesANPH Midterm NotesClaudine Jo B. TalabocNo ratings yet
- 3D. Muscle TissueDocument47 pages3D. Muscle Tissuejaf123xyNo ratings yet
- 589bc774 Df1d 47c6 Bc97 B5fd73030ca6muscle PhysiologyDocument3 pages589bc774 Df1d 47c6 Bc97 B5fd73030ca6muscle PhysiologyMalti SharmaNo ratings yet
- Skeletal MusclesDocument7 pagesSkeletal MusclesAhmed OudahNo ratings yet
- Muscular SystemDocument6 pagesMuscular SystemTricia Mae LlerenaNo ratings yet
- Locomotion & Movement (Muscles)Document41 pagesLocomotion & Movement (Muscles)DEEPANKAR SAHANo ratings yet
- Excitable Tissues 1Document8 pagesExcitable Tissues 1gojaisaiah787No ratings yet
- Locomotion and Movement Notes Grade 11Document15 pagesLocomotion and Movement Notes Grade 11brahmatej99No ratings yet
- Types of Muscle TissueDocument11 pagesTypes of Muscle TissueAndrea Louise ReyesNo ratings yet
- B Muscle Tissue HistologyDocument55 pagesB Muscle Tissue HistologyAffie SaikolNo ratings yet
- Muscular SystemDocument12 pagesMuscular SystemChrisjohn Aster OsioNo ratings yet
- Muscle PhysiologyDocument50 pagesMuscle PhysiologyJoyce Adjei-boateng100% (1)
- Muscle Tissue OutlineDocument4 pagesMuscle Tissue OutlinePeachy PieNo ratings yet
- Anaphy-Muscular HandoutDocument7 pagesAnaphy-Muscular HandoutAEGEL DARO TORRALBANo ratings yet
- ST - of Myofilaments..Document10 pagesST - of Myofilaments..Mohammed HamadaNo ratings yet
- Muscle Tissue LectureDocument34 pagesMuscle Tissue LectureurjaNo ratings yet
- Muscle TissueDocument11 pagesMuscle TissueAlexNo ratings yet
- Muscle: Drags Them inDocument5 pagesMuscle: Drags Them inKhaldun MirNo ratings yet
- Reviewer - Muscular SystemDocument11 pagesReviewer - Muscular SystemIvy Jan OcateNo ratings yet
- Topic 7 Muscular SystemDocument7 pagesTopic 7 Muscular SystemMary Ann Morillo TenederoNo ratings yet
- MUSC System WuDocument65 pagesMUSC System Wuhayatseid2002No ratings yet
- Chapter 7 - Muscular SystemDocument29 pagesChapter 7 - Muscular SystemlNo ratings yet
- SDL MuscularDocument6 pagesSDL MuscularMonique Eloise GualizaNo ratings yet
- Lecture 8 (Muscular System Lecture 2)Document43 pagesLecture 8 (Muscular System Lecture 2)Husnain WattoNo ratings yet
- Myosin Powered MovementDocument8 pagesMyosin Powered MovementIndranil BanerjeeNo ratings yet
- Faculty of Medicine: Practical Work No. 9Document3 pagesFaculty of Medicine: Practical Work No. 9Andrei TimofteNo ratings yet
- MUSCLE and NERVOUSHISTOLOGYDocument78 pagesMUSCLE and NERVOUSHISTOLOGYAerielle De GuzmanNo ratings yet
- HAPP Week VII XDocument32 pagesHAPP Week VII Xxuxi dulNo ratings yet
- Human Anatomy and Physiology - Skeletal MuscleDocument14 pagesHuman Anatomy and Physiology - Skeletal MuscleRAI SUNNo ratings yet
- Sliding Filament Theory of Muscle Contraction: Muscular TissueDocument8 pagesSliding Filament Theory of Muscle Contraction: Muscular TissueMaryam Nur Azizah RahmahNo ratings yet
- Locomotion and MovementDocument9 pagesLocomotion and Movementpoojari.dedeepyaNo ratings yet
- Skeletal Muscle Contraction and The Motor UnitsDocument4 pagesSkeletal Muscle Contraction and The Motor UnitsAnonymous vnv6QFNo ratings yet
- Skeletal Muscle StructureDocument38 pagesSkeletal Muscle Structuretoyy100% (1)
- Muscle TissueDocument31 pagesMuscle Tissueaimi BatrisyiaNo ratings yet
- Muscle Tissue NotesDocument11 pagesMuscle Tissue NotesAubrey Unique EvangelistaNo ratings yet
- Skeletal MuscleDocument3 pagesSkeletal Musclekrystal1994No ratings yet
- Muscle PhysiologyDocument24 pagesMuscle PhysiologyArmando AlehandroNo ratings yet
- Wa0038Document10 pagesWa0038Shehzad QureshiNo ratings yet
- MusclesssDocument4 pagesMusclesssLiam Jacque LapuzNo ratings yet
- Human Muscular System: Types of Movement in Human BeingDocument4 pagesHuman Muscular System: Types of Movement in Human Beingaadeshthite476No ratings yet
- M223 Ancestral Journey Eng PublDocument28 pagesM223 Ancestral Journey Eng PublGábor BaloghNo ratings yet
- Güntürkün, Bugnyar (2016) Cognition Without CortexDocument13 pagesGüntürkün, Bugnyar (2016) Cognition Without CortexMak PavičevićNo ratings yet
- Exercise Bio IV-Plantae Part 1Document1 pageExercise Bio IV-Plantae Part 1Sella OktavianiNo ratings yet
- Cryptosporidium en AguasDocument138 pagesCryptosporidium en AguasJennyfer Carolina Alejo RiverosNo ratings yet
- Rheumatoid ArthritisDocument12 pagesRheumatoid Arthritissri subhikshaNo ratings yet
- Act 8 - Corn Chi Square AnalysisDocument5 pagesAct 8 - Corn Chi Square AnalysisChristine PazNo ratings yet
- 3 Enzymes Paper 2 - N-MQDocument67 pages3 Enzymes Paper 2 - N-MQMostafa AmerNo ratings yet
- Growth HormoneDocument7 pagesGrowth Hormonefs semjohn100% (1)
- Brachionus CalyciflorusDocument10 pagesBrachionus CalyciflorusÉrica OliveiraNo ratings yet
- Biology Investigatory Project Class 12 PDFDocument35 pagesBiology Investigatory Project Class 12 PDFJubilee sharma0% (2)
- The Deaths and Side Effects of Toxic Covid Vaccines!Document56 pagesThe Deaths and Side Effects of Toxic Covid Vaccines!Francisco100% (1)
- Second Year Botany Ipe Importatn QuesitonsDocument4 pagesSecond Year Botany Ipe Importatn Quesitonsj72jpjcjsbNo ratings yet
- Desordenes Del Desarrollo Sexual Copia 2Document16 pagesDesordenes Del Desarrollo Sexual Copia 2mariakjaimesNo ratings yet
- Replication ErrorsDocument16 pagesReplication Errorsilkay turanNo ratings yet
- Rivera-Laboratory Exercise No. 2-AnaphyDocument3 pagesRivera-Laboratory Exercise No. 2-AnaphyRivera, Trishia PilonNo ratings yet
- Bio 10 Master Study Guide Part 1Document9 pagesBio 10 Master Study Guide Part 1leonor.estimaNo ratings yet
- GMM. WHO CNS5 Classification (English Version)Document8 pagesGMM. WHO CNS5 Classification (English Version)Melissa Garcia LezamaNo ratings yet
- Endocrine System: Cell Secretion Types Types of HormonesDocument6 pagesEndocrine System: Cell Secretion Types Types of HormonesKrissia BaasisNo ratings yet
- 2.6 Structure of DNA & RNADocument3 pages2.6 Structure of DNA & RNATeena Tutor100% (1)
- II Puc Biology Practicals - Mid TermDocument7 pagesII Puc Biology Practicals - Mid TermSatish GaNo ratings yet
- Cell Organelles and FunctionsDocument2 pagesCell Organelles and Functionsyurie mendozaNo ratings yet
- Running Head: STEM CELL BIOLOGY 1Document4 pagesRunning Head: STEM CELL BIOLOGY 1alexNo ratings yet
- A Century of Hardy-Weinberg EquilibriumDocument9 pagesA Century of Hardy-Weinberg EquilibriumPriawanIndraNo ratings yet
- Jurnal Biomedika: Patricia Gita NaullyDocument6 pagesJurnal Biomedika: Patricia Gita NaullyAchmad Nur AlamNo ratings yet
- 5090 s13 QP 11Document16 pages5090 s13 QP 11Khaled MohammadNo ratings yet
- Sci8 Q4 Mod6Document30 pagesSci8 Q4 Mod6Alessa LamesNo ratings yet
- Biology CP Study Guide (DNA, RNA, & Protein Synthesis) Answer KeyDocument4 pagesBiology CP Study Guide (DNA, RNA, & Protein Synthesis) Answer Keymisterbrowner100% (2)
- Do Jews Carry Trauma in Our Genes - David Samuels (Articulo)Document14 pagesDo Jews Carry Trauma in Our Genes - David Samuels (Articulo)Gustavo BenaderetNo ratings yet
- © Lesson Plans Inc. 2007Document9 pages© Lesson Plans Inc. 2007Fatima Nur Faiza HandaNo ratings yet












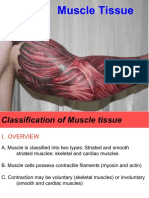







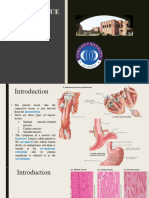













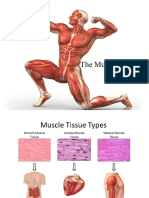












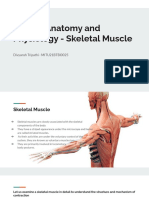










































Download as pptx, pdf, or txt
You might also like
- Biomolecules Recap Answer Key by The Amoeba Sisters AnswerkeyDocument3 pagesBiomolecules Recap Answer Key by The Amoeba Sisters AnswerkeySandra Lopez50% (2)
- Muscular TissueDocument65 pagesMuscular TissueMitzel SapaloNo ratings yet
- Histology of MuscleDocument49 pagesHistology of MuscleTewachew Dessie100% (1)
- Muscle Tissue-Wl06.11Document54 pagesMuscle Tissue-Wl06.11api-19641337No ratings yet
- 29.-31. SBRC MSK SummaryDocument153 pages29.-31. SBRC MSK SummaryErin HillNo ratings yet
- Chap-20 - Locomotion & MovementDocument52 pagesChap-20 - Locomotion & MovementMittal SavaniNo ratings yet
- Muscular TDocument18 pagesMuscular TSumayya KabeerNo ratings yet
- MascularDocument39 pagesMascularAive BelistaNo ratings yet
- Muscle Tissue 2012Document13 pagesMuscle Tissue 2012ST. Fatimah KadirNo ratings yet
- FF Raumenys PDFDocument34 pagesFF Raumenys PDFLaura PaškevičiūtėNo ratings yet
- Muscle Tissue Lesson 1Document5 pagesMuscle Tissue Lesson 1Steves LeeboyNo ratings yet
- Muscle Tissue: - Skeletal Muscle - Cardiac Muscle - Smooth MuscleDocument36 pagesMuscle Tissue: - Skeletal Muscle - Cardiac Muscle - Smooth MuscleDentist Dina Samy100% (1)
- Muscular TissuesDocument46 pagesMuscular TissuesDAVE CANALETANo ratings yet
- Sweeney 2018Document14 pagesSweeney 2018LuisJauweriNo ratings yet
- 6.muscular TissueDocument40 pages6.muscular Tissuemero wowNo ratings yet
- 04muscle TissueDocument39 pages04muscle Tissueapi-19916399No ratings yet
- Histology of Cardiac and Smooth MuscleDocument73 pagesHistology of Cardiac and Smooth MuscleShehryar AbbasNo ratings yet
- Histology of Cardiac and Smooth MuscleDocument73 pagesHistology of Cardiac and Smooth MuscleShehryar AbbasNo ratings yet
- LPT 321Document103 pagesLPT 321rahul roshanNo ratings yet
- ANPH Midterm NotesDocument44 pagesANPH Midterm NotesClaudine Jo B. TalabocNo ratings yet
- 3D. Muscle TissueDocument47 pages3D. Muscle Tissuejaf123xyNo ratings yet
- 589bc774 Df1d 47c6 Bc97 B5fd73030ca6muscle PhysiologyDocument3 pages589bc774 Df1d 47c6 Bc97 B5fd73030ca6muscle PhysiologyMalti SharmaNo ratings yet
- Skeletal MusclesDocument7 pagesSkeletal MusclesAhmed OudahNo ratings yet
- Muscular SystemDocument6 pagesMuscular SystemTricia Mae LlerenaNo ratings yet
- Locomotion & Movement (Muscles)Document41 pagesLocomotion & Movement (Muscles)DEEPANKAR SAHANo ratings yet
- Excitable Tissues 1Document8 pagesExcitable Tissues 1gojaisaiah787No ratings yet
- Locomotion and Movement Notes Grade 11Document15 pagesLocomotion and Movement Notes Grade 11brahmatej99No ratings yet
- Types of Muscle TissueDocument11 pagesTypes of Muscle TissueAndrea Louise ReyesNo ratings yet
- B Muscle Tissue HistologyDocument55 pagesB Muscle Tissue HistologyAffie SaikolNo ratings yet
- Muscular SystemDocument12 pagesMuscular SystemChrisjohn Aster OsioNo ratings yet
- Muscle PhysiologyDocument50 pagesMuscle PhysiologyJoyce Adjei-boateng100% (1)
- Muscle Tissue OutlineDocument4 pagesMuscle Tissue OutlinePeachy PieNo ratings yet
- Anaphy-Muscular HandoutDocument7 pagesAnaphy-Muscular HandoutAEGEL DARO TORRALBANo ratings yet
- ST - of Myofilaments..Document10 pagesST - of Myofilaments..Mohammed HamadaNo ratings yet
- Muscle Tissue LectureDocument34 pagesMuscle Tissue LectureurjaNo ratings yet
- Muscle TissueDocument11 pagesMuscle TissueAlexNo ratings yet
- Muscle: Drags Them inDocument5 pagesMuscle: Drags Them inKhaldun MirNo ratings yet
- Reviewer - Muscular SystemDocument11 pagesReviewer - Muscular SystemIvy Jan OcateNo ratings yet
- Topic 7 Muscular SystemDocument7 pagesTopic 7 Muscular SystemMary Ann Morillo TenederoNo ratings yet
- MUSC System WuDocument65 pagesMUSC System Wuhayatseid2002No ratings yet
- Chapter 7 - Muscular SystemDocument29 pagesChapter 7 - Muscular SystemlNo ratings yet
- SDL MuscularDocument6 pagesSDL MuscularMonique Eloise GualizaNo ratings yet
- Lecture 8 (Muscular System Lecture 2)Document43 pagesLecture 8 (Muscular System Lecture 2)Husnain WattoNo ratings yet
- Myosin Powered MovementDocument8 pagesMyosin Powered MovementIndranil BanerjeeNo ratings yet
- Faculty of Medicine: Practical Work No. 9Document3 pagesFaculty of Medicine: Practical Work No. 9Andrei TimofteNo ratings yet
- MUSCLE and NERVOUSHISTOLOGYDocument78 pagesMUSCLE and NERVOUSHISTOLOGYAerielle De GuzmanNo ratings yet
- HAPP Week VII XDocument32 pagesHAPP Week VII Xxuxi dulNo ratings yet
- Human Anatomy and Physiology - Skeletal MuscleDocument14 pagesHuman Anatomy and Physiology - Skeletal MuscleRAI SUNNo ratings yet
- Sliding Filament Theory of Muscle Contraction: Muscular TissueDocument8 pagesSliding Filament Theory of Muscle Contraction: Muscular TissueMaryam Nur Azizah RahmahNo ratings yet
- Locomotion and MovementDocument9 pagesLocomotion and Movementpoojari.dedeepyaNo ratings yet
- Skeletal Muscle Contraction and The Motor UnitsDocument4 pagesSkeletal Muscle Contraction and The Motor UnitsAnonymous vnv6QFNo ratings yet
- Skeletal Muscle StructureDocument38 pagesSkeletal Muscle Structuretoyy100% (1)
- Muscle TissueDocument31 pagesMuscle Tissueaimi BatrisyiaNo ratings yet
- Muscle Tissue NotesDocument11 pagesMuscle Tissue NotesAubrey Unique EvangelistaNo ratings yet
- Skeletal MuscleDocument3 pagesSkeletal Musclekrystal1994No ratings yet
- Muscle PhysiologyDocument24 pagesMuscle PhysiologyArmando AlehandroNo ratings yet
- Wa0038Document10 pagesWa0038Shehzad QureshiNo ratings yet
- MusclesssDocument4 pagesMusclesssLiam Jacque LapuzNo ratings yet
- Human Muscular System: Types of Movement in Human BeingDocument4 pagesHuman Muscular System: Types of Movement in Human Beingaadeshthite476No ratings yet
- M223 Ancestral Journey Eng PublDocument28 pagesM223 Ancestral Journey Eng PublGábor BaloghNo ratings yet
- Güntürkün, Bugnyar (2016) Cognition Without CortexDocument13 pagesGüntürkün, Bugnyar (2016) Cognition Without CortexMak PavičevićNo ratings yet
- Exercise Bio IV-Plantae Part 1Document1 pageExercise Bio IV-Plantae Part 1Sella OktavianiNo ratings yet
- Cryptosporidium en AguasDocument138 pagesCryptosporidium en AguasJennyfer Carolina Alejo RiverosNo ratings yet
- Rheumatoid ArthritisDocument12 pagesRheumatoid Arthritissri subhikshaNo ratings yet
- Act 8 - Corn Chi Square AnalysisDocument5 pagesAct 8 - Corn Chi Square AnalysisChristine PazNo ratings yet
- 3 Enzymes Paper 2 - N-MQDocument67 pages3 Enzymes Paper 2 - N-MQMostafa AmerNo ratings yet
- Growth HormoneDocument7 pagesGrowth Hormonefs semjohn100% (1)
- Brachionus CalyciflorusDocument10 pagesBrachionus CalyciflorusÉrica OliveiraNo ratings yet
- Biology Investigatory Project Class 12 PDFDocument35 pagesBiology Investigatory Project Class 12 PDFJubilee sharma0% (2)
- The Deaths and Side Effects of Toxic Covid Vaccines!Document56 pagesThe Deaths and Side Effects of Toxic Covid Vaccines!Francisco100% (1)
- Second Year Botany Ipe Importatn QuesitonsDocument4 pagesSecond Year Botany Ipe Importatn Quesitonsj72jpjcjsbNo ratings yet
- Desordenes Del Desarrollo Sexual Copia 2Document16 pagesDesordenes Del Desarrollo Sexual Copia 2mariakjaimesNo ratings yet
- Replication ErrorsDocument16 pagesReplication Errorsilkay turanNo ratings yet
- Rivera-Laboratory Exercise No. 2-AnaphyDocument3 pagesRivera-Laboratory Exercise No. 2-AnaphyRivera, Trishia PilonNo ratings yet
- Bio 10 Master Study Guide Part 1Document9 pagesBio 10 Master Study Guide Part 1leonor.estimaNo ratings yet
- GMM. WHO CNS5 Classification (English Version)Document8 pagesGMM. WHO CNS5 Classification (English Version)Melissa Garcia LezamaNo ratings yet
- Endocrine System: Cell Secretion Types Types of HormonesDocument6 pagesEndocrine System: Cell Secretion Types Types of HormonesKrissia BaasisNo ratings yet
- 2.6 Structure of DNA & RNADocument3 pages2.6 Structure of DNA & RNATeena Tutor100% (1)
- II Puc Biology Practicals - Mid TermDocument7 pagesII Puc Biology Practicals - Mid TermSatish GaNo ratings yet
- Cell Organelles and FunctionsDocument2 pagesCell Organelles and Functionsyurie mendozaNo ratings yet
- Running Head: STEM CELL BIOLOGY 1Document4 pagesRunning Head: STEM CELL BIOLOGY 1alexNo ratings yet
- A Century of Hardy-Weinberg EquilibriumDocument9 pagesA Century of Hardy-Weinberg EquilibriumPriawanIndraNo ratings yet
- Jurnal Biomedika: Patricia Gita NaullyDocument6 pagesJurnal Biomedika: Patricia Gita NaullyAchmad Nur AlamNo ratings yet
- 5090 s13 QP 11Document16 pages5090 s13 QP 11Khaled MohammadNo ratings yet
- Sci8 Q4 Mod6Document30 pagesSci8 Q4 Mod6Alessa LamesNo ratings yet
- Biology CP Study Guide (DNA, RNA, & Protein Synthesis) Answer KeyDocument4 pagesBiology CP Study Guide (DNA, RNA, & Protein Synthesis) Answer Keymisterbrowner100% (2)
- Do Jews Carry Trauma in Our Genes - David Samuels (Articulo)Document14 pagesDo Jews Carry Trauma in Our Genes - David Samuels (Articulo)Gustavo BenaderetNo ratings yet
- © Lesson Plans Inc. 2007Document9 pages© Lesson Plans Inc. 2007Fatima Nur Faiza HandaNo ratings yet