Download as pptx, pdf, or txt
You might also like
- Preclinical Physiology Review 2023: For USMLE Step 1 and COMLEX-USA Level 1From EverandPreclinical Physiology Review 2023: For USMLE Step 1 and COMLEX-USA Level 1Rating: 1 out of 5 stars1/5 (1)
- The Ammonicarb Operation Book Format 4 PDF FreeDocument149 pagesThe Ammonicarb Operation Book Format 4 PDF FreePantelis Krintz100% (1)
- Biochem ABO IncompatibilityDocument62 pagesBiochem ABO IncompatibilityChi Samaniego100% (4)
- Membrane DeformabilityDocument2 pagesMembrane DeformabilityKenneth Jake BatiduanNo ratings yet
- RBC MembraneDocument41 pagesRBC MembranemwambungupNo ratings yet
- Erythrocyte Metabolism AND Membrane Structure and FunctionDocument29 pagesErythrocyte Metabolism AND Membrane Structure and FunctionJohn Marie IdavaNo ratings yet
- Red Blood Cell BiochemistryDocument19 pagesRed Blood Cell BiochemistryPrincewill SeiyefaNo ratings yet
- Plasma Membrane: The Lipid Bilayer Membrane ProteinsDocument79 pagesPlasma Membrane: The Lipid Bilayer Membrane ProteinsEarfanNo ratings yet
- Lipid Movement BiochemDocument9 pagesLipid Movement BiochemCrowNo ratings yet
- G Rojos BlancosDocument7 pagesG Rojos BlancosAndrea Valenzuela AedoNo ratings yet
- Module 3 - Lecture 3 - LectureDocument8 pagesModule 3 - Lecture 3 - LectureNi CaoNo ratings yet
- RBC MembraneDocument37 pagesRBC MembraneStanley ChikoveNo ratings yet
- 01 Cell MembraneDocument22 pages01 Cell MembraneMian SahibNo ratings yet
- Chapter 9 HemaDocument4 pagesChapter 9 HemaJoannah MarieNo ratings yet
- Study Exercise Biology 10-12 97.03Document7 pagesStudy Exercise Biology 10-12 97.03Fabiola Vania FeliciaNo ratings yet
- Research Paper On Plasma MembraneDocument4 pagesResearch Paper On Plasma Membranevvgnzdbkf100% (1)
- Erythrocyte Metabolism and Membrane Structure and FunctionDocument38 pagesErythrocyte Metabolism and Membrane Structure and FunctionRej MaeNo ratings yet
- Chapter 4-Jnu Biomembranes-1 - 複本Document48 pagesChapter 4-Jnu Biomembranes-1 - 複本Wai Kwong ChiuNo ratings yet
- Components of Plasma Membranes: Learning ObjectivesDocument14 pagesComponents of Plasma Membranes: Learning ObjectivesAlthea Junsan BernabeNo ratings yet
- Biochemistry & Genetics: II SHS 109: Resource Person: DR Tanveer Akbar Reference TextDocument60 pagesBiochemistry & Genetics: II SHS 109: Resource Person: DR Tanveer Akbar Reference TexttNo ratings yet
- The Cell MembraneDocument36 pagesThe Cell Membraneanulorance98No ratings yet
- Cell MembraneDocument2 pagesCell Membranehello123No ratings yet
- Lect 1 Membrane Structure and FunctionDocument67 pagesLect 1 Membrane Structure and FunctionIzukanji kayoraNo ratings yet
- Topic 3Document12 pagesTopic 3Ria AccadNo ratings yet
- Life at The Edge Concept 7.1: Structure! Cellular Membranes Are Fluid Mosaics of Lipids and ProteinsDocument6 pagesLife at The Edge Concept 7.1: Structure! Cellular Membranes Are Fluid Mosaics of Lipids and ProteinsAisha100% (1)
- Plasma Membrane Research PapersDocument6 pagesPlasma Membrane Research Papersaflbasnka100% (1)
- Lipid Traffic Floppy Drives and A SuperhighwayDocument12 pagesLipid Traffic Floppy Drives and A SuperhighwayJohan AlarcónNo ratings yet
- Red Blood Cell MembraneDocument28 pagesRed Blood Cell MembraneAisyah Atikah100% (1)
- Plasma Membrane in DetailsDocument24 pagesPlasma Membrane in DetailsSyeda Kaneez Mohammadi MohammadiNo ratings yet
- CellDocument48 pagesCellSoumik SahaNo ratings yet
- Kuliah MembranDocument61 pagesKuliah Membrannikmah nuur rochmahNo ratings yet
- Solution BiotechDocument54 pagesSolution Biotechkavita sahaiNo ratings yet
- Biological Cell MembranesDocument44 pagesBiological Cell MembranesMohammed ShaikNo ratings yet
- Chemistry of Lipid Domains: Advanced Article Richard M. EpandDocument5 pagesChemistry of Lipid Domains: Advanced Article Richard M. EpandazzaassNo ratings yet
- Chapter 5 Lipids Membrane LectureslidesDocument65 pagesChapter 5 Lipids Membrane Lectureslidesduy trầnNo ratings yet
- Chapter Blood: RBC Platelet HemostasisDocument89 pagesChapter Blood: RBC Platelet Hemostasisapi-19916399100% (1)
- Plasma MembraneDocument35 pagesPlasma MembraneMini RůžičkaNo ratings yet
- Cell Membrane (Structure and Function)Document150 pagesCell Membrane (Structure and Function)Dr. Tapan Kr. Dutta100% (1)
- CellMembrane Structure and Functional PropertiesDocument5 pagesCellMembrane Structure and Functional PropertiesAnastasiafynnNo ratings yet
- K - 8 Blood Brain Barrier (Biokimia)Document31 pagesK - 8 Blood Brain Barrier (Biokimia)Firman YunusNo ratings yet
- Biochemistry ProblemDocument5 pagesBiochemistry ProblemJnanendra SenguptaNo ratings yet
- Red Cell Membrane Structure 2Document16 pagesRed Cell Membrane Structure 2MONo ratings yet
- Fisiologi Cairan Tubuh & Darah: Dr. Nindya Aryanty, M.Med - EdDocument43 pagesFisiologi Cairan Tubuh & Darah: Dr. Nindya Aryanty, M.Med - EdIman TaufiqNo ratings yet
- Introductory One Written Answers: 1) Mention 4 Sultar Containing GAGS and The Function of Two of ThemDocument8 pagesIntroductory One Written Answers: 1) Mention 4 Sultar Containing GAGS and The Function of Two of ThemEbrahim RagabNo ratings yet
- Plasma MembraneDocument18 pagesPlasma MembraneJemuel Bucud LagartoNo ratings yet
- Lesson 3 Red Cell Memebrane Developing-Metabolic PathwaysDocument19 pagesLesson 3 Red Cell Memebrane Developing-Metabolic Pathwaysتجربة أولىNo ratings yet
- Membrane Structure and Function: Lecture OutlineDocument11 pagesMembrane Structure and Function: Lecture Outlinehaha_le12No ratings yet
- Normal Erythropoiesis: A. Inner and Outer LayersDocument9 pagesNormal Erythropoiesis: A. Inner and Outer LayersTina MultazamiNo ratings yet
- Biochem ReportDocument10 pagesBiochem ReportLahari PylaNo ratings yet
- Unit 2 ReviewerDocument4 pagesUnit 2 Revieweraaleah moscaNo ratings yet
- Composition of Blood DR - Fayaz MemonDocument29 pagesComposition of Blood DR - Fayaz Memonhifzaaijaz366No ratings yet
- Lecture 4 - MembraneDocument30 pagesLecture 4 - MembranePhạm ĐứcNo ratings yet
- Lecture 4 - MembraneDocument30 pagesLecture 4 - MembranePhạm ĐứcNo ratings yet
- 03lect Cell2 Summary2017Document4 pages03lect Cell2 Summary2017Dan ChoiNo ratings yet
- Plasma Membrane and Transport MechanismsDocument63 pagesPlasma Membrane and Transport MechanismsVaccedNo ratings yet
- Chemical Composition of Blood Cells: MLS 302: Basic HaematologyDocument13 pagesChemical Composition of Blood Cells: MLS 302: Basic HaematologyBarakat IsmailNo ratings yet
- Rbcs Metabolism: D.Hythem SaeedDocument19 pagesRbcs Metabolism: D.Hythem Saeedsamar yousif mohamedNo ratings yet
- Zoology Lecture 4Document18 pagesZoology Lecture 4Stephen OcheaNo ratings yet
- HUBS2206 - Human Biochemistry and CellDocument87 pagesHUBS2206 - Human Biochemistry and CellLuke ThomsonNo ratings yet
- X Membrane and TransportDocument35 pagesX Membrane and TransportHitesh MoreNo ratings yet
- Red Blood Cells, Functions, Diseases A Simple Guide To The Condition, Diagnosis, Treatment, And Related ConditionsFrom EverandRed Blood Cells, Functions, Diseases A Simple Guide To The Condition, Diagnosis, Treatment, And Related ConditionsNo ratings yet
- Activity 4-Smearing and Simple StainingDocument7 pagesActivity 4-Smearing and Simple StainingMyedelle SeacorNo ratings yet
- Governmental Health AgenciesDocument4 pagesGovernmental Health AgenciesMyedelle SeacorNo ratings yet
- 6 ETS VenipunctureDocument7 pages6 ETS VenipunctureMyedelle SeacorNo ratings yet
- Phlebotomy TechniqueDocument2 pagesPhlebotomy TechniqueMyedelle SeacorNo ratings yet
- The Philippines During Martial Law: "Suko Na! Talo Ka Na! Marcos, LAYAS NA!" - Sigaw NG BayanDocument23 pagesThe Philippines During Martial Law: "Suko Na! Talo Ka Na! Marcos, LAYAS NA!" - Sigaw NG BayanMyedelle SeacorNo ratings yet
- MLS109Lab LabApp&MicroscopeDocument4 pagesMLS109Lab LabApp&MicroscopeMyedelle SeacorNo ratings yet
- Phase of MatterDocument12 pagesPhase of MatterMyedelle SeacorNo ratings yet
- Physical Characteristics: Functions of BloodDocument5 pagesPhysical Characteristics: Functions of BloodMyedelle SeacorNo ratings yet
- Normoblastic Mat Urat IonDocument15 pagesNormoblastic Mat Urat IonMyedelle SeacorNo ratings yet
- Medical Examiner Qualifications: Please PRINT ClearlyDocument2 pagesMedical Examiner Qualifications: Please PRINT ClearlyMyedelle SeacorNo ratings yet
- Transwomen Are Women: Writing PromptDocument2 pagesTranswomen Are Women: Writing PromptMyedelle SeacorNo ratings yet
- The Phil Blood Bank Laws-PART1Document5 pagesThe Phil Blood Bank Laws-PART1Myedelle SeacorNo ratings yet
- REFERENCE NUMBER: 000014120: and Mathematics (Stem)Document2 pagesREFERENCE NUMBER: 000014120: and Mathematics (Stem)Myedelle SeacorNo ratings yet
- How To Write A Position PaperDocument10 pagesHow To Write A Position PaperMyedelle SeacorNo ratings yet
- The Phil Blood Bank Laws-PART1Document6 pagesThe Phil Blood Bank Laws-PART1Myedelle SeacorNo ratings yet
- Empowerment Technologies Midterms ReviewerDocument11 pagesEmpowerment Technologies Midterms ReviewerMyedelle SeacorNo ratings yet
- 12-Stem-C Capstone Research Project: Name Field Capstone TitleDocument1 page12-Stem-C Capstone Research Project: Name Field Capstone TitleMyedelle SeacorNo ratings yet
- Sts History ReviewerDocument3 pagesSts History ReviewerMyedelle SeacorNo ratings yet
- Introduction To The LessonDocument4 pagesIntroduction To The LessonMyedelle SeacorNo ratings yet
- Chemistry Investigatory ProjectDocument16 pagesChemistry Investigatory ProjectDibyanshu NathNo ratings yet
- Asm1 21200Document6 pagesAsm1 21200nooneNo ratings yet
- Human Anatomy and Physiology 2nd Edition Amerman Test BankDocument32 pagesHuman Anatomy and Physiology 2nd Edition Amerman Test BankallisontaylorfnrqzamgksNo ratings yet
- Pgurl 2679036134142300Document4 pagesPgurl 2679036134142300Barbara VasquezNo ratings yet
- Major Classification of Indian SoilsDocument4 pagesMajor Classification of Indian SoilsALABANI MANGANo ratings yet
- Collagen StructureDocument42 pagesCollagen StructureAatmaanandaNo ratings yet
- Carbohydrates NotesDocument28 pagesCarbohydrates Noteschaitanyaa baliNo ratings yet
- Pig Iron Blast Furnace: Mcqs Preparation For Engineering Competitive ExamsDocument20 pagesPig Iron Blast Furnace: Mcqs Preparation For Engineering Competitive ExamschauhanNo ratings yet
- Lec 27Document18 pagesLec 27capcanNo ratings yet
- Hard Gelatin CapsulesDocument58 pagesHard Gelatin Capsulesas.jessy0608No ratings yet
- Ufi-String D PDFDocument4 pagesUfi-String D PDFNermeen ElmelegaeNo ratings yet
- 3 LAB ACTIVITY SHEET 3 Precipitation and Solubility 1Document3 pages3 LAB ACTIVITY SHEET 3 Precipitation and Solubility 1Jurel JohnNo ratings yet
- Chemistry Content Based ExpirimentsDocument22 pagesChemistry Content Based ExpirimentsHari VaarthanNo ratings yet
- Dowex™ Monosphere™ 88: Product Type Matrix Functional GroupDocument2 pagesDowex™ Monosphere™ 88: Product Type Matrix Functional GroupBrianNo ratings yet
- Class 12 Chemistry MCQ Worksheet-2Document7 pagesClass 12 Chemistry MCQ Worksheet-28D audio StationNo ratings yet
- Chemistry of BORON and SILICONDocument32 pagesChemistry of BORON and SILICONprashantNo ratings yet
- MSDSDocument25 pagesMSDShenry.anthirayasNo ratings yet
- HDRscholrarship AustraliaDocument2 pagesHDRscholrarship AustraliaMuhammad Sohail AshrafNo ratings yet
- Preparation of ColloidsDocument11 pagesPreparation of ColloidsNurhidayah Mat Sarit80% (5)
- Preparation of Cupramonium Rayon ThreadDocument22 pagesPreparation of Cupramonium Rayon ThreadRahul MNo ratings yet
- Textbook of CosmetologyDocument96 pagesTextbook of CosmetologyRicky SetiawanNo ratings yet
- CLASS VIII QUESTION BANK - 4. Metals and Non-MetalsDocument7 pagesCLASS VIII QUESTION BANK - 4. Metals and Non-MetalsSurbhi NayarNo ratings yet
- Advanced Concrete Technology Past Paper Questions 1Document11 pagesAdvanced Concrete Technology Past Paper Questions 1Asad QaziNo ratings yet
- 56 102 1 SMDocument6 pages56 102 1 SMAzka Hamam BaihaqiNo ratings yet
- Keerthi M.Nair, Vignesh Kumaravel, Suresh C Pillai Carbonaceous Cathode Materials For Electro-Fenton TechnologyDocument142 pagesKeerthi M.Nair, Vignesh Kumaravel, Suresh C Pillai Carbonaceous Cathode Materials For Electro-Fenton TechnologyVăn Thành MaiNo ratings yet
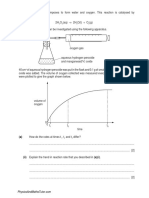
- 1 Hydrogen Peroxide Decomposes To Form Water and Oxygen. This Reaction Is Catalysed byDocument9 pages1 Hydrogen Peroxide Decomposes To Form Water and Oxygen. This Reaction Is Catalysed byKelvin DenhereNo ratings yet
- and 68070 S2012 Final v2 No AnswersDocument42 pagesand 68070 S2012 Final v2 No AnswersZadrin TuckerNo ratings yet
- 2016 Book TheZincBromineFlowBatteryDocument113 pages2016 Book TheZincBromineFlowBatteryraghu7862No ratings yet
- A Combined Spectroscopy and Computational MoleculaDocument13 pagesA Combined Spectroscopy and Computational MoleculadubasilioNo ratings yet