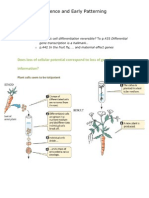
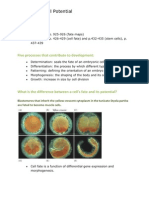
Download as ppt, pdf, or txt
You might also like
- Chime Sog Thig Part 1 - Shenphen Dawa RinpocheDocument45 pagesChime Sog Thig Part 1 - Shenphen Dawa Rinpochesusco100% (4)
- Extranuclear Inheritance-ExamplesDocument18 pagesExtranuclear Inheritance-ExamplesTony BernardNo ratings yet
- Csir Unit 5 Updated PDFDocument10 pagesCsir Unit 5 Updated PDFAmaldev S K100% (2)
- Lorenzo L. Taping III BS BiologyDocument60 pagesLorenzo L. Taping III BS BiologyEnzo TapingNo ratings yet
- Role of Cytoplasm in Cell DifferentiationDocument7 pagesRole of Cytoplasm in Cell DifferentiationJ LALNo ratings yet
- Organogenesis and Morphogenesis in PlantsDocument51 pagesOrganogenesis and Morphogenesis in PlantsElgie JimenezNo ratings yet
- Morphogenetic Mechanisms1Document36 pagesMorphogenetic Mechanisms1Lily VaughNo ratings yet
- 203 RibetDocument9 pages203 RibetSohan PkNo ratings yet
- Teacher: DR Gouri Course: Zoology Hons. 6 Sem Paper: Developmental BiologyDocument17 pagesTeacher: DR Gouri Course: Zoology Hons. 6 Sem Paper: Developmental BiologyApratim SinghNo ratings yet
- C. Elegans Fertilization and CleavageDocument3 pagesC. Elegans Fertilization and CleavageJanine VillaflorNo ratings yet
- C ElegansDocument10 pagesC ElegansMmMmGoodNo ratings yet
- Cleavage in MammalsDocument51 pagesCleavage in MammalsIrfan Azram100% (4)
- All Sexual Organisms Begin As A Single CellDocument12 pagesAll Sexual Organisms Begin As A Single CellAFRIADMA AULIA PERDANANo ratings yet
- Differential Gene Expression and DevelopmentDocument9 pagesDifferential Gene Expression and DevelopmentShaher Bano Mirza100% (1)
- Genomic Equivalence and Early PatterningDocument11 pagesGenomic Equivalence and Early PatterningSavannah Simone PetrachenkoNo ratings yet
- Extra Nuclear InheritanceDocument13 pagesExtra Nuclear InheritanceTony BernardNo ratings yet
- Hematopoiesis Stem CellsDocument9 pagesHematopoiesis Stem CellsslxuNo ratings yet
- BIO125ZusammenfassungLF 3Document43 pagesBIO125ZusammenfassungLF 399rnbccdr8No ratings yet
- Somatic Embryogenesis PDFDocument6 pagesSomatic Embryogenesis PDFeduardo9119No ratings yet
- 2009 Midterm Introduction To Life Science I. Q&A (70%) : BMES - Chuang CYDocument8 pages2009 Midterm Introduction To Life Science I. Q&A (70%) : BMES - Chuang CYHsieh Chia YuNo ratings yet
- 7 Gendev PDFDocument12 pages7 Gendev PDFJohn DeHansNo ratings yet
- Mitosis and Meiosis Pre-WorksheetDocument5 pagesMitosis and Meiosis Pre-WorksheetRoshae SinclairNo ratings yet
- John J Harada: Signaling in Plant EmbryogenesisDocument5 pagesJohn J Harada: Signaling in Plant Embryogenesisshady182No ratings yet
- Aleurone LayerDocument5 pagesAleurone LayerMian RashidNo ratings yet
- Pembelahan Embrio (Cleavage) S2 2017Document47 pagesPembelahan Embrio (Cleavage) S2 2017Desy NataliaNo ratings yet
- Inv 13Document9 pagesInv 13mrojasf.1996No ratings yet
- BIO109P Module 2. Differentiation & MorphogenesisDocument28 pagesBIO109P Module 2. Differentiation & MorphogenesisJet DelimaNo ratings yet
- Molecular Regulation of Pancreatic Beta-Cell Mass DevelopmentDocument14 pagesMolecular Regulation of Pancreatic Beta-Cell Mass DevelopmentAndrei MitescuNo ratings yet
- Cell Fate and Cell PotentialDocument12 pagesCell Fate and Cell PotentialSavannah Simone PetrachenkoNo ratings yet
- M06 Axisspecificationindrosophila Quad 1Document18 pagesM06 Axisspecificationindrosophila Quad 1Mahendra KatreNo ratings yet
- GeneticsDocument4 pagesGeneticsrizamaevillanueva79No ratings yet
- Ectopic Trancript of BBMDocument13 pagesEctopic Trancript of BBMMalik NADA-ABINo ratings yet
- Biochemistry of Embryogenesis in MonocotsDocument11 pagesBiochemistry of Embryogenesis in MonocotsAmrit Kumar NayakNo ratings yet
- Heterogeneity in Oct4 and Sox2 Targets Biases CellDocument15 pagesHeterogeneity in Oct4 and Sox2 Targets Biases CellJoy SahaNo ratings yet
- Unit 16 Mechanisms of Cell Interaction: StructureDocument25 pagesUnit 16 Mechanisms of Cell Interaction: Structurekaladhar reddyNo ratings yet
- Asymmetrical Cell: Submitted byDocument5 pagesAsymmetrical Cell: Submitted byJeffrey VictoryNo ratings yet
- Testis Development: Juho-Antti Mäkelä, Jaakko J. Koskenniemi, Helena E. Virtanen, and Jorma ToppariDocument49 pagesTestis Development: Juho-Antti Mäkelä, Jaakko J. Koskenniemi, Helena E. Virtanen, and Jorma TopparigüldenNo ratings yet
- Activity 1Document2 pagesActivity 1Jes RenNo ratings yet
- BIO Mod2 Cell Growth and ReproductionDocument10 pagesBIO Mod2 Cell Growth and Reproductiongeorge linNo ratings yet
- Unit 2 QuestionsDocument3 pagesUnit 2 QuestionssseettNo ratings yet
- Garces 2007Document6 pagesGarces 2007Peter CordovageNo ratings yet
- Biochemistry Digital Assignment: by S. Manoj Kumar 18BAG0106Document7 pagesBiochemistry Digital Assignment: by S. Manoj Kumar 18BAG0106Ãppu ŠrkNo ratings yet
- BIO 325 Exam 1 ReviewDocument17 pagesBIO 325 Exam 1 ReviewNerdy Notes Inc.No ratings yet
- Apoptosis Study SheetDocument7 pagesApoptosis Study SheetAshima SarupNo ratings yet
- Meiosis: Dr. Shayma'a J. Ahmed Prof. Genetic Engineerin& BiotechnologyDocument31 pagesMeiosis: Dr. Shayma'a J. Ahmed Prof. Genetic Engineerin& BiotechnologyMariam QaisNo ratings yet
- Cell LineageDocument7 pagesCell LineageAmar Kant JhaNo ratings yet
- Drill 2 CELL CYCLEDocument4 pagesDrill 2 CELL CYCLEJoyce FernandezNo ratings yet
- Genes Development FinalDocument59 pagesGenes Development FinalJennifer ValdezNo ratings yet
- Green Beginnings - Pattern Formation in The Early Plant EmbryoDocument27 pagesGreen Beginnings - Pattern Formation in The Early Plant EmbryoMark DenhamNo ratings yet
- Lab 9Document40 pagesLab 9bob0% (1)
- 0000 2005 Nature NSC Cell and NeurogenesisDocument12 pages0000 2005 Nature NSC Cell and NeurogenesisEsteban Daniel Peluffo ArgónNo ratings yet
- The Big Mitosis and Meiosis Quiz-1Document12 pagesThe Big Mitosis and Meiosis Quiz-1api-246382283No ratings yet
- Module 2 Cell Cycle and Cell DivisionDocument9 pagesModule 2 Cell Cycle and Cell DivisionBalolot RalphNo ratings yet
- Biology 9700 Notes Topic 16 Inherited Change 2019-20Document32 pagesBiology 9700 Notes Topic 16 Inherited Change 2019-20ADEEL AHMAD67% (3)
- (Medical Journal of Cell Biology) Cytoplasmic and Nuclear Maturation of Oocytes in Mammals Living in The Shadow of Cells Developmental CapabilityDocument5 pages(Medical Journal of Cell Biology) Cytoplasmic and Nuclear Maturation of Oocytes in Mammals Living in The Shadow of Cells Developmental CapabilityBagas KarangzNo ratings yet
- AP Biology Lab 3 QuestionsDocument3 pagesAP Biology Lab 3 Questionsedwarddame100% (1)
- Biology Module 2010 SeratasDocument28 pagesBiology Module 2010 SeratasNurul AzuwinNo ratings yet
- Ezrin Becomes Restricted To Outer Cells Following Asymmetrical Division in The Preimplantation Mouse EmbryoDocument12 pagesEzrin Becomes Restricted To Outer Cells Following Asymmetrical Division in The Preimplantation Mouse EmbryonugrahoneyNo ratings yet
- Biology Unit 16Document14 pagesBiology Unit 16joyNo ratings yet
- 1 s2.0 S0012160620303195 Main 1 PDFDocument20 pages1 s2.0 S0012160620303195 Main 1 PDFNabila NajwaNo ratings yet
- Data and VariableDocument12 pagesData and VariableAlamgir HossainNo ratings yet
- Biostatistics: Written by - Alomgir HossainDocument7 pagesBiostatistics: Written by - Alomgir HossainAlamgir HossainNo ratings yet
- Structured Association Mapping Using STRDocument15 pagesStructured Association Mapping Using STRAlamgir HossainNo ratings yet
- SSR PrimersDocument53 pagesSSR PrimersAlamgir HossainNo ratings yet
- Assignment On MycobacteriaDocument7 pagesAssignment On MycobacteriaAlamgir HossainNo ratings yet
- Biostatistics CourseDocument100 pagesBiostatistics CourseAlamgir Hossain100% (1)
- Microbiology''', Is The Study of Microbes, Such AsDocument27 pagesMicrobiology''', Is The Study of Microbes, Such AsAlamgir HossainNo ratings yet
- Animal Breeding GEB 206Document21 pagesAnimal Breeding GEB 206Alamgir HossainNo ratings yet
- Coefficient Variation - Anil SirDocument5 pagesCoefficient Variation - Anil SirAlamgir HossainNo ratings yet
- Bone DevelopmentDocument26 pagesBone DevelopmentAlamgir HossainNo ratings yet
- Jabsco Marine Product Guide 2006.Document24 pagesJabsco Marine Product Guide 2006.m_najmanNo ratings yet
- INTRODUCTION of Digital RecruitmentDocument11 pagesINTRODUCTION of Digital Recruitmentpawan bathamNo ratings yet
- Chapter 6 Hexavalent ChromiumDocument56 pagesChapter 6 Hexavalent Chromiumgabriel100% (1)
- CBSE Class 9 Science Revision Notes Chapter - 1 Matter in Our SurroundingsDocument68 pagesCBSE Class 9 Science Revision Notes Chapter - 1 Matter in Our SurroundingsParesh RanjanNo ratings yet
- 2.3.5 Practice - Atomic Structure (Practice)Document11 pages2.3.5 Practice - Atomic Structure (Practice)damarionbryant46No ratings yet
- Mountain Man Brewery Company Case AnalysisDocument4 pagesMountain Man Brewery Company Case AnalysisAmit PathakNo ratings yet
- Chen 2021Document19 pagesChen 2021saka healthNo ratings yet
- Arcelormittal Galati S.A. - Romania: Inspection Certificate 20564275 According To: en 10204/2004/3.1 Date: 26.11.2018Document1 pageArcelormittal Galati S.A. - Romania: Inspection Certificate 20564275 According To: en 10204/2004/3.1 Date: 26.11.2018Felicia Cioaba100% (2)
- Letter of Undertaking Letter Seggregation in ChargeDocument1 pageLetter of Undertaking Letter Seggregation in ChargeSijin SivankuttyNo ratings yet
- 2011 Douglas County Fair - Schedule of EventsDocument3 pages2011 Douglas County Fair - Schedule of EventsNews-Review of Roseburg OregonNo ratings yet
- Sol-Gel Methods PDFDocument19 pagesSol-Gel Methods PDFDimitrios Tsiplakides100% (2)
- CPG - Management of AcneDocument97 pagesCPG - Management of Acneumiraihana1No ratings yet
- Heat TransferDocument27 pagesHeat TransferLurking RogueNo ratings yet
- Fact Sheet On The Death of Rey RiveraDocument4 pagesFact Sheet On The Death of Rey Riverahillarymitchell_5921No ratings yet
- Pembagian Distribusi Publikasi Peserta Poster IcprpDocument8 pagesPembagian Distribusi Publikasi Peserta Poster IcprpimaNo ratings yet
- Digitalhospital AnexampleofbestpracticeDocument8 pagesDigitalhospital AnexampleofbestpracticeTesfa Eu DegefeyNo ratings yet
- Basic Waste Management Training After Revision (Compatibility Mode)Document64 pagesBasic Waste Management Training After Revision (Compatibility Mode)Gangsar LukmanjayaNo ratings yet
- Types of TaxesDocument15 pagesTypes of TaxesNischal KumarNo ratings yet
- ETP-295 SISTEMAS DE RECUBRIMIENTOS ANTICORROSIVOS E IDENTIFICACIÓN PARA INSTALACIONES SUPERFICIALES - EnglishDocument25 pagesETP-295 SISTEMAS DE RECUBRIMIENTOS ANTICORROSIVOS E IDENTIFICACIÓN PARA INSTALACIONES SUPERFICIALES - EnglishArcadio Duran100% (1)
- LC GCDocument23 pagesLC GCEsra'A Subhi Sabri QasemNo ratings yet
- A Rasch Analysis On Collapsing Categories in Item - S Response Scales of Survey Questionnaire Maybe It - S Not One Size Fits All PDFDocument29 pagesA Rasch Analysis On Collapsing Categories in Item - S Response Scales of Survey Questionnaire Maybe It - S Not One Size Fits All PDFNancy NgNo ratings yet
- Delayed-Action AcceleratorDocument3 pagesDelayed-Action AcceleratorMOHD FIRDAUSNo ratings yet
- Mobile Crane SafetyDocument3 pagesMobile Crane SafetyAnonymous LFgO4WbID0% (1)
- Lipid Analysis: An Introduction ToDocument8 pagesLipid Analysis: An Introduction ToElaiza HerreraNo ratings yet
- Polyaluminium Chloride MSDS DikonversiDocument7 pagesPolyaluminium Chloride MSDS DikonversiPUJI ASTUTINo ratings yet
- 2-322 Physical and Chemical Data: X Y X YDocument9 pages2-322 Physical and Chemical Data: X Y X YKelo CastestNo ratings yet
- Nora (2019) Quick and Easy Vegan Family Dinners EBOOKDocument45 pagesNora (2019) Quick and Easy Vegan Family Dinners EBOOKShawn A. Wygant100% (2)
- Process and Material Specification: AIPI05-03-006 01-02-71 A1 FEB 01/21Document17 pagesProcess and Material Specification: AIPI05-03-006 01-02-71 A1 FEB 01/21schembriryanNo ratings yet