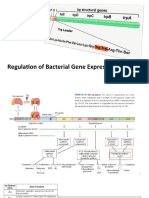
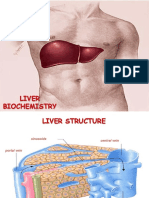
Download as pptx, pdf, or txt
You might also like
- TRP OperonDocument5 pagesTRP OperonrahulhaldankarNo ratings yet
- Regulation of Gene Expression in ProkaryotesDocument4 pagesRegulation of Gene Expression in ProkaryotesBelay Rayos del SolNo ratings yet
- Lecture - 1Document33 pagesLecture - 1Bahesty Monfared AkashNo ratings yet
- The TRP OperonDocument7 pagesThe TRP OperonnikhilsathwikNo ratings yet
- Lecturtry Operon 3Document4 pagesLecturtry Operon 3maxwell amponsahNo ratings yet
- FUNCTION AND CONTROL OF THE TRYPTOPHAN TRPDocument23 pagesFUNCTION AND CONTROL OF THE TRYPTOPHAN TRPkizzaymenteraNo ratings yet
- Advanced Genetics Q2Document1 pageAdvanced Genetics Q2Mohd Amirul Firdhaus Mohd RidhwanNo ratings yet
- 2017-18 - T-Genetica Debacterias e Virus-P4Document13 pages2017-18 - T-Genetica Debacterias e Virus-P4João DomingosNo ratings yet
- Gene Regulation: Mrs. Ofelia Solano SaludarDocument39 pagesGene Regulation: Mrs. Ofelia Solano SaludarmskikiNo ratings yet
- Translation (Biology)Document22 pagesTranslation (Biology)Manoj JoshiNo ratings yet
- TRP Operon PresentationDocument27 pagesTRP Operon PresentationPradeep SinghNo ratings yet
- Wa0008.Document17 pagesWa0008.Rita JaypuriaNo ratings yet
- Attenuation, Control of Gene Expression By: Advanced ArticleDocument12 pagesAttenuation, Control of Gene Expression By: Advanced ArticleazzaassNo ratings yet
- Gene RegulationDocument86 pagesGene RegulationMeenal MeshramNo ratings yet
- Translation 30th March 2009Document16 pagesTranslation 30th March 2009api-26794308No ratings yet
- TRANSLATIONDocument19 pagesTRANSLATIONUsha SuthersonNo ratings yet
- Gene RegulationDocument4 pagesGene RegulationPetros MaheyaNo ratings yet
- Translation 190603145457Document30 pagesTranslation 190603145457soniaazizNo ratings yet
- Kul Biomol Bakteri Ekspresi GenDocument28 pagesKul Biomol Bakteri Ekspresi GenERIE YUWITA SARINo ratings yet
- CH 2E - Gene RegulationDocument19 pagesCH 2E - Gene Regulationtanush2007sekharNo ratings yet
- TRANSLATIONDocument12 pagesTRANSLATIONNEET STUDIESNo ratings yet
- Transcription &translation: Mol. Biology Lec-4 TranscriptionDocument12 pagesTranscription &translation: Mol. Biology Lec-4 TranscriptionAhmed Ali AssafNo ratings yet
- Gene Expression in ProkaryotesDocument43 pagesGene Expression in ProkaryotesravibiriNo ratings yet
- Lec. 6Document10 pagesLec. 6Dr. Mohamed ShamsNo ratings yet
- Regulation of Histidine and Hut OperonsDocument11 pagesRegulation of Histidine and Hut Operonsaditi_joshee419No ratings yet
- Bio Short Notes FinalDocument5 pagesBio Short Notes FinalMUHAMMAD HAFIZ FAZRISHAH BIN JEFFRY KIMNo ratings yet
- Utkarsh Sharma XII - A Biology Investigatory Project - TranslationDocument24 pagesUtkarsh Sharma XII - A Biology Investigatory Project - TranslationUtkarsh SharmaNo ratings yet
- Biochemistry: RNA Synthesis and ProcessingDocument54 pagesBiochemistry: RNA Synthesis and ProcessingAqsa YaminNo ratings yet
- Meredith Barb Homework-2 (25 Points) Due On Sept. 24 by 5:00pmDocument5 pagesMeredith Barb Homework-2 (25 Points) Due On Sept. 24 by 5:00pmMeredith BarbNo ratings yet
- Translation and Regulation of Gene ExpressionDocument51 pagesTranslation and Regulation of Gene ExpressionP. Jacksen Sam PaulNo ratings yet
- Translation AnoverviewDocument32 pagesTranslation AnoverviewshitalchandrasitNo ratings yet
- 6 Regulation of Gene ExpressionDocument49 pages6 Regulation of Gene ExpressionNaivedyam Vineeta MishraNo ratings yet
- Transcription in EukaryotesDocument49 pagesTranscription in EukaryotesmrlabdesigneNo ratings yet
- Chromosomal Map Begins at Oric Units of MinutesDocument10 pagesChromosomal Map Begins at Oric Units of Minutesdhita malikaNo ratings yet
- Regulation of Gene Expression in Eukaryotes: Presented by Quratulain (19-Arid-1403) Asadullah (19-Arid-1379)Document32 pagesRegulation of Gene Expression in Eukaryotes: Presented by Quratulain (19-Arid-1403) Asadullah (19-Arid-1379)Beauty LiciousNo ratings yet
- Protein Synthesis: Transcription & TranslationDocument4 pagesProtein Synthesis: Transcription & TranslationtshahriyarNo ratings yet
- Control of Gene ExpressionDocument9 pagesControl of Gene Expressionaloysius limNo ratings yet
- 07 Control of Gene Expression in ProkaryotesDocument17 pages07 Control of Gene Expression in Prokaryotesmustafa aliNo ratings yet
- Try Operon 2Document25 pagesTry Operon 2maxwell amponsahNo ratings yet
- Biochem AssDocument9 pagesBiochem AssBereket AntenehNo ratings yet
- Transcriptiontranslation 170210171328Document34 pagesTranscriptiontranslation 170210171328Farah B. BtoushNo ratings yet
- (123doc) - Operons-Va-Qua-Trinh-Dieu-Hoa-Hoat-Dong-Cua-GenDocument24 pages(123doc) - Operons-Va-Qua-Trinh-Dieu-Hoa-Hoat-Dong-Cua-GenKhanh VyNo ratings yet
- Gene ExpressionDocument58 pagesGene ExpressionJunirose PanesNo ratings yet
- Euk TranscriptionDocument22 pagesEuk TranscriptionManish DasNo ratings yet
- Lecture 3Document6 pagesLecture 3tiaria.wilson24No ratings yet
- Transcription in ProkaryotesDocument18 pagesTranscription in ProkaryotesPrakash100% (1)
- Genetics DocsDocument2 pagesGenetics Docsjayarbenaning1No ratings yet
- Transcription Prokaryotes 2012-cDocument27 pagesTranscription Prokaryotes 2012-cAnupama PatiNo ratings yet
- Gene Regulation in Prokaryotes-1Document17 pagesGene Regulation in Prokaryotes-1Fasiha Mushadi100% (1)
- Gene ExpressionDocument19 pagesGene Expressionkvicto100% (1)
- Central Dogma: Faculty of Medicine, Sriwijaya UniversityDocument51 pagesCentral Dogma: Faculty of Medicine, Sriwijaya UniversityIlsya PertiwiNo ratings yet
- Replication - Transcription - TranslationDocument75 pagesReplication - Transcription - TranslationJason FryNo ratings yet
- Protein SynthesisDocument135 pagesProtein SynthesisCarlaNo ratings yet
- Messenger Rnas (Mrnas) : This Class of Rnas Are The Genetic Coding TemplatesDocument8 pagesMessenger Rnas (Mrnas) : This Class of Rnas Are The Genetic Coding TemplatesRavi AlugubelliNo ratings yet
- Translation Bio Ch6Document12 pagesTranslation Bio Ch6Hordit LyngdohNo ratings yet
- Transcription: Protein Biosynthesis Is The Process by Which BiologicalDocument4 pagesTranscription: Protein Biosynthesis Is The Process by Which BiologicalAnand RajNo ratings yet
- TRANSLATION - PicsDocument62 pagesTRANSLATION - PicsJisna JaisonNo ratings yet
- Otot 2Document106 pagesOtot 2anita parwatiNo ratings yet
- Molecular Final1Document9 pagesMolecular Final1h4hh9wqwvnNo ratings yet
- Final Dna Damage and RepairDocument32 pagesFinal Dna Damage and RepairARUN KUMARNo ratings yet
- Site Directed MutagenesisDocument22 pagesSite Directed MutagenesisARUN KUMARNo ratings yet
- Post-Transcriptional Modifications: Arun Kumar PHD Scholar Centre For Molecular Biology Central University of JammuDocument67 pagesPost-Transcriptional Modifications: Arun Kumar PHD Scholar Centre For Molecular Biology Central University of JammuARUN KUMARNo ratings yet
- Next Generation SequencingDocument70 pagesNext Generation SequencingARUN KUMARNo ratings yet
- Introduction To Biotechnology - Chapter 1Document54 pagesIntroduction To Biotechnology - Chapter 1sisil safitriNo ratings yet
- Digestion Lab CauseyDocument3 pagesDigestion Lab CauseynicewanNo ratings yet
- Molecular Diversity - WheatDocument8 pagesMolecular Diversity - WheatHarshvardhan ZalaNo ratings yet
- Metabolisme Kolesterol, Lipoprotein Dan ApolipoproteinDocument32 pagesMetabolisme Kolesterol, Lipoprotein Dan ApolipoproteinAlmira Ulfa Utari NasutionNo ratings yet
- IB Biology Notes - 36 EnzymesDocument2 pagesIB Biology Notes - 36 EnzymesJohn Philip D. NapalNo ratings yet
- Physiology, Lecture 3 (Pictures Only)Document40 pagesPhysiology, Lecture 3 (Pictures Only)Ali Al-QudsiNo ratings yet
- Atomi HSC Biology Study Plan 2023Document2 pagesAtomi HSC Biology Study Plan 2023Emanuel YakoNo ratings yet
- Mbt1 B Cloning Expression 14 15Document91 pagesMbt1 B Cloning Expression 14 15Abhishek SinghNo ratings yet
- AA Protein EnzymeDocument64 pagesAA Protein EnzymeNash DeniegaNo ratings yet
- ProteinsDocument11 pagesProteinsGabrielle AmadorNo ratings yet
- University of Alkafeel Faculty of Medical & Health Techniques Dept. of Medical Laboratory TechniquesDocument3 pagesUniversity of Alkafeel Faculty of Medical & Health Techniques Dept. of Medical Laboratory Techniquesحسن الأسديNo ratings yet
- Biochem Exam Practice QuestionsDocument12 pagesBiochem Exam Practice Questionsihack_101No ratings yet
- Anaerobic Energy MetabolismDocument1 pageAnaerobic Energy MetabolismSoham DibyachintanNo ratings yet
- Final Burtons SummaryDocument53 pagesFinal Burtons SummaryJessabelle Ramos100% (1)
- IBO 2010 Korea Theory Answers 2Document30 pagesIBO 2010 Korea Theory Answers 2martynapetNo ratings yet
- Multigenre Project - Teaching Cellular Respiration and PhotosynthesisDocument11 pagesMultigenre Project - Teaching Cellular Respiration and Photosynthesisapi-398387480No ratings yet
- Biol 110 Essex Without Recitation Spring 2015Document7 pagesBiol 110 Essex Without Recitation Spring 2015api-281761748No ratings yet
- Promega Catalog 2020 Ca030Document408 pagesPromega Catalog 2020 Ca030Stefy TorresNo ratings yet
- DNA TranscriptionDocument1 pageDNA TranscriptionJanelle sarmientoNo ratings yet
- Dipolar Coupling: RDCS, Structure, Dynamics and Disorder! Embo Course 2013!Document59 pagesDipolar Coupling: RDCS, Structure, Dynamics and Disorder! Embo Course 2013!Rigel_TNo ratings yet
- 26th Biosciences Quiz Bee ReviewerDocument12 pages26th Biosciences Quiz Bee ReviewerEuan Joshua B. Evangelista100% (1)
- Liver and CurcuminDocument6 pagesLiver and CurcuminLouie Mae SantosNo ratings yet
- Water, PH and BuffersDocument43 pagesWater, PH and BuffersDaniel LuchendoNo ratings yet
- HIV and AIDSDocument75 pagesHIV and AIDSMarc Imhotep Cray, M.D.100% (1)
- Draft Sooca Case 1 2014Document22 pagesDraft Sooca Case 1 2014Hafdzi MaulanaNo ratings yet
- Gradien, 10 (2), Pp. 992-995Document3 pagesGradien, 10 (2), Pp. 992-995Riza Ma'rufinNo ratings yet
- Biochemistry and Cell Biology of Ageing: Part III Biomedical ScienceDocument424 pagesBiochemistry and Cell Biology of Ageing: Part III Biomedical ScienceAureliano BuendiaNo ratings yet
- Lesson Discussion: Self-Learning Module (SLM) General Biology 2Document7 pagesLesson Discussion: Self-Learning Module (SLM) General Biology 2almafebe caselNo ratings yet
- Gastroenterology Clinics PDFDocument211 pagesGastroenterology Clinics PDFjackNo ratings yet
- Biochemical Functions of LiverDocument94 pagesBiochemical Functions of LiverMi PatelNo ratings yet