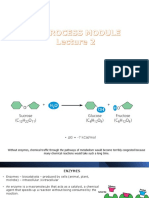
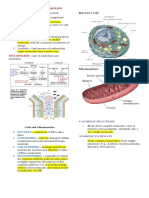
Download as ppt, pdf, or txt
You might also like
- Effect of PH On The Activity of Potato CatalaseDocument1 pageEffect of PH On The Activity of Potato CatalaseNathuAndrews50% (2)
- 1FFF11B1BD16627EE05400144FEB5F70.pptDocument63 pages1FFF11B1BD16627EE05400144FEB5F70.pptNur AishaNo ratings yet
- Module 2 Enzyme TransDocument11 pagesModule 2 Enzyme TransKathleen CedilloNo ratings yet
- EnzymeDocument86 pagesEnzymeFiraol DiribaNo ratings yet
- Lecture 2Document18 pagesLecture 2warden tambiNo ratings yet
- 4 EnzymesDocument33 pages4 EnzymesmjmonforteNo ratings yet
- Biological Chemistry: Enzyme Kinetics Part 1Document30 pagesBiological Chemistry: Enzyme Kinetics Part 1Mohammed shaffiqueNo ratings yet
- ENZYMESDocument55 pagesENZYMESAsante BrightNo ratings yet
- Biochem NursingDocument144 pagesBiochem Nursingabukaritoyibu100No ratings yet
- Enzymes NotesDocument46 pagesEnzymes NotesCuthbert MweembaNo ratings yet
- Enzymes LectureDocument120 pagesEnzymes LectureHERSEY MIAYONo ratings yet
- Enzymes PDFDocument37 pagesEnzymes PDFOmer KareemNo ratings yet
- Enzymes - Part IIDocument71 pagesEnzymes - Part IIBarış KaplanNo ratings yet
- Lecture 1 - The Kinetics of Enzyme Catalyzed ReactionsDocument18 pagesLecture 1 - The Kinetics of Enzyme Catalyzed ReactionsGenevive S. de VeraNo ratings yet
- Biochemistry of Enzymes & Clinical Enzymology: By: Tesfahun MollaDocument156 pagesBiochemistry of Enzymes & Clinical Enzymology: By: Tesfahun MollaAddis MémñøňNo ratings yet
- EnzymesDocument58 pagesEnzymesFateh RaufNo ratings yet
- Module 4 - Enzymes 1 PDFDocument14 pagesModule 4 - Enzymes 1 PDFFrancis ValdezNo ratings yet
- Enzyme Catalysis, Types of Mechanisms Involved.: David Enoma 17PCP0163 1Document18 pagesEnzyme Catalysis, Types of Mechanisms Involved.: David Enoma 17PCP0163 1Vijay Kumar ReddyNo ratings yet
- Materi 2Document34 pagesMateri 2siti purnamaNo ratings yet
- Enzymes LectureDocument115 pagesEnzymes LectureJane Tai100% (1)
- 9744 H2 Biology Lecture Notes - JC 1: EnzymesDocument43 pages9744 H2 Biology Lecture Notes - JC 1: EnzymesTimothy HandokoNo ratings yet
- Lecture 2 Enzyme KineticsDocument31 pagesLecture 2 Enzyme KineticsCarinaJongLeeNo ratings yet
- Unit 2.1 Clinic EnzymologyDocument26 pagesUnit 2.1 Clinic EnzymologyGuillermo LasarteNo ratings yet
- Medical Biochemistry: Enzymes: Mr. Genaro F. Alderite JR, MsermDocument58 pagesMedical Biochemistry: Enzymes: Mr. Genaro F. Alderite JR, MsermNash DeniegaNo ratings yet
- Study On EnzymesDocument21 pagesStudy On EnzymesPragyan Kumar PradhanNo ratings yet
- Enzymes: Bettelheim / Brown / Campbell / Farrell / TorresDocument51 pagesEnzymes: Bettelheim / Brown / Campbell / Farrell / TorresMica BernardoNo ratings yet
- Biochemistry: By: Angela Marie Ferrer BSN 2BDocument13 pagesBiochemistry: By: Angela Marie Ferrer BSN 2BNoemi Martinez FerrerNo ratings yet
- Enzymes and Biological: CatalysisDocument41 pagesEnzymes and Biological: CatalysisKiya AlemuNo ratings yet
- Chapter 23Document53 pagesChapter 23haririNo ratings yet
- Enzyme KineticsDocument10 pagesEnzyme Kineticskedirwoliy100% (1)
- Lecture 9 PDFDocument24 pagesLecture 9 PDFNazir KhanNo ratings yet
- Biological Molecules: Prepared By: Mrs. Eden C. SanchezDocument50 pagesBiological Molecules: Prepared By: Mrs. Eden C. SanchezBernard D. Fajardo Jr.100% (1)
- Introduction To Enzymes and Mechanism of Enzyme ActionDocument40 pagesIntroduction To Enzymes and Mechanism of Enzyme ActionAtif Amin BaigNo ratings yet
- Enzymes and Their RegulationDocument48 pagesEnzymes and Their RegulationChathur Chandrasoorya100% (1)
- BT1000 - Cellular Metabolism PDFDocument35 pagesBT1000 - Cellular Metabolism PDFAnubhavAgarwal100% (1)
- Enzymes Kinetics and Specificity: 生物化學一 林重銘老師 AA509 EXT 3955 第 版Document72 pagesEnzymes Kinetics and Specificity: 生物化學一 林重銘老師 AA509 EXT 3955 第 版陳家琪No ratings yet
- ICBI121 Lecture5 MetabolismDocument60 pagesICBI121 Lecture5 MetabolismSupasinee RNo ratings yet
- EnzymesDocument3 pagesEnzymesArabela SimanganNo ratings yet
- Midterm Biochem ReviewerDocument19 pagesMidterm Biochem ReviewerNATALIE NICOLE GABASNo ratings yet
- Enzyme Kinetics and Applications (Part 1a: Kinetics of Enzyme Catalyzed Reactions)Document26 pagesEnzyme Kinetics and Applications (Part 1a: Kinetics of Enzyme Catalyzed Reactions)Nur AishaNo ratings yet
- Untitled Document 1Document9 pagesUntitled Document 1Zwen Zyronne Norico LumiwesNo ratings yet
- 1 Bioenegretics and MetabolismDocument60 pages1 Bioenegretics and MetabolismlikeleliletabaNo ratings yet
- Industrial Microbiology W5Document97 pagesIndustrial Microbiology W5Yaren ErelNo ratings yet
- EnzymeDocument15 pagesEnzymeSheena Mhae LopeñaNo ratings yet
- Stocker Enzymes and VitaminsDocument75 pagesStocker Enzymes and VitaminsDylan SalesNo ratings yet
- Enzyme 1Document115 pagesEnzyme 1fikaduNo ratings yet
- Enzymes BKSDocument74 pagesEnzymes BKSAjmain ShakibNo ratings yet
- 3 EnzymesDocument27 pages3 Enzymesharshit khareNo ratings yet
- ENZYMESDocument31 pagesENZYMESjuunisai6No ratings yet
- 5 - CHEM 46 - Chapter 5 EnzymesDocument50 pages5 - CHEM 46 - Chapter 5 EnzymesbeautybaylantacsimbajonNo ratings yet
- LEC 06 - Enzyme MechanismsDocument70 pagesLEC 06 - Enzyme Mechanismseliza makNo ratings yet
- Chapter 5 CatalysisDocument31 pagesChapter 5 CatalysisHien HoangNo ratings yet
- Enzyme: Ns. Shila Wisnasari, S.Kep., M.BiomedDocument55 pagesEnzyme: Ns. Shila Wisnasari, S.Kep., M.BiomedSyahdaJuvenilProfitamelaNo ratings yet
- 02 EnzymesDocument63 pages02 EnzymesFatish BanguraNo ratings yet
- Voet Chapt 11Document112 pagesVoet Chapt 11Sandhya KanetkarNo ratings yet
- Key Suggestions For MBBS StudentsDocument14 pagesKey Suggestions For MBBS Studentsnihadabj2003No ratings yet
- Prelab 4Document6 pagesPrelab 4Trần Xuân QuỳnhNo ratings yet
- Enzymes PDFDocument36 pagesEnzymes PDFAltamashNo ratings yet
- EnzymesDocument35 pagesEnzymesRicardo Cubillos UribeNo ratings yet
- Summary Biology IB High Level Topic 8Document21 pagesSummary Biology IB High Level Topic 8Magdalena CzajaNo ratings yet
- Biotransfrmtns Prepartv Organic Chemistry: The Use of Isolated Enzymes and Whole Cell Systems in SynthesisFrom EverandBiotransfrmtns Prepartv Organic Chemistry: The Use of Isolated Enzymes and Whole Cell Systems in SynthesisNo ratings yet
- Enzyme KineticsDocument90 pagesEnzyme KineticsAmrit LalNo ratings yet
- Practice Exam HSN104 ANSWERSDocument13 pagesPractice Exam HSN104 ANSWERSRaha RajabigamasaeiNo ratings yet
- PL - ProDia ChemistryDocument4 pagesPL - ProDia ChemistryChristian SenopadangNo ratings yet
- Chapter 4Document47 pagesChapter 4api-19968742No ratings yet
- BIOMOLECULESDocument54 pagesBIOMOLECULESTanmayNo ratings yet
- F3 Assignment 3.1 - Balanced DietDocument2 pagesF3 Assignment 3.1 - Balanced DietAshleyNo ratings yet
- Nutrients and Food TestingDocument17 pagesNutrients and Food TestingThanigaivelan A/L Arumugam [Student]No ratings yet
- Transfer RNA Synthesis and Regulation: Advanced ArticleDocument17 pagesTransfer RNA Synthesis and Regulation: Advanced ArticleAfif BerlianNo ratings yet
- Replication ErrorsDocument16 pagesReplication Errorsilkay turanNo ratings yet
- Structural Homology Modeling of C-Terminal Domain of The Dystrophin Protein: An In-Silico ApproachDocument8 pagesStructural Homology Modeling of C-Terminal Domain of The Dystrophin Protein: An In-Silico ApproachInternational Journal of Innovative Science and Research Technology100% (1)
- Enzyme SpecificityDocument1 pageEnzyme SpecificityEarl FredNo ratings yet
- Enzymes A Practical Introduction To Structure Mechanism and Data Analysis 3Rd Edition Robert A Copeland Full ChapterDocument62 pagesEnzymes A Practical Introduction To Structure Mechanism and Data Analysis 3Rd Edition Robert A Copeland Full Chapterkevin.smart540100% (7)
- General Zoology Module 1 Lesson 3Document3 pagesGeneral Zoology Module 1 Lesson 3Paul PanizaNo ratings yet
- LIPIDSDocument30 pagesLIPIDSMary Ann OgoyNo ratings yet
- Glycogen MetabolismDocument11 pagesGlycogen MetabolismShruti MishraNo ratings yet
- Sarcoplasmic Reticulum 3Document6 pagesSarcoplasmic Reticulum 3Syaidah umiNo ratings yet
- IB BIOLOGY SL TOPIC 3 Nucleic Acids & Proteins and Chemical Elements & WaterDocument13 pagesIB BIOLOGY SL TOPIC 3 Nucleic Acids & Proteins and Chemical Elements & WaterweeNo ratings yet
- Lectura 9 Benefits of Organic EggsDocument2 pagesLectura 9 Benefits of Organic EggsDavid moraNo ratings yet
- 4.3met Karbohidrat (Fileminimizer)Document63 pages4.3met Karbohidrat (Fileminimizer)Hendra SudaryonoNo ratings yet
- Nutrition, Metabolism, and Body Temperature RegulationDocument11 pagesNutrition, Metabolism, and Body Temperature RegulationSNo ratings yet
- SBIA031 Study Guide 2024Document10 pagesSBIA031 Study Guide 2024Itumeleng SefaraNo ratings yet
- Qualitative Analysis of Amino Acids and Proteins and Carbohydrates and Lipids and SteroidsDocument35 pagesQualitative Analysis of Amino Acids and Proteins and Carbohydrates and Lipids and SteroidsgangsNo ratings yet
- DNA Damage and RepairDocument32 pagesDNA Damage and RepairNashmiya SaheerNo ratings yet
- Performance Task #1 - Aceron, Barrina, VelasDocument6 pagesPerformance Task #1 - Aceron, Barrina, VelasCalil AceronNo ratings yet
- Biomolecules Chemistry AssignmentDocument28 pagesBiomolecules Chemistry AssignmentAnurag Singh Thakur67% (3)
- I. Objectives: Lesson Plan in Grade 9Document3 pagesI. Objectives: Lesson Plan in Grade 9Maria Karina Ferreras100% (1)
- NCERT Class 12 Chemistry BiomoleculesDocument22 pagesNCERT Class 12 Chemistry BiomoleculesAnant MalhotraNo ratings yet
- Nad / Nadp: 3. Coenzyme Q / UbiquinoneDocument4 pagesNad / Nadp: 3. Coenzyme Q / UbiquinoneNicole ChanNo ratings yet
- Amino Acids: Central University of Himachal PradeshDocument22 pagesAmino Acids: Central University of Himachal PradeshSaurav Bhardwaj100% (1)