Download as pptx, pdf, or txt
You might also like
- ULTRASOUND Abdomen Normal MeasurementsDocument20 pagesULTRASOUND Abdomen Normal Measurementsadinatop100% (4)
- Cardiovascular System: Mungcal, Dharlynette RTRPDocument77 pagesCardiovascular System: Mungcal, Dharlynette RTRPDharlyn MungcalNo ratings yet
- Visceral Vascular Manipulation, 2011Document270 pagesVisceral Vascular Manipulation, 2011kkalu_just_funky91% (11)
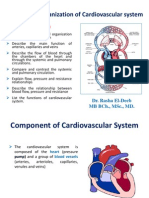
- Functional Organization of Cardiovascular SystemDocument19 pagesFunctional Organization of Cardiovascular SystemIbtesam Mohammed100% (2)
- Anatomy and Physiology of The Cardiovascular SystemDocument6 pagesAnatomy and Physiology of The Cardiovascular SystemDawn Santiago RNNo ratings yet
- Anatomy and Physiology by Ross and WilsonDocument21 pagesAnatomy and Physiology by Ross and WilsonAnonymous iEjePNZ50% (2)
- Circulatory System: A Tutorial Study GuideFrom EverandCirculatory System: A Tutorial Study GuideRating: 5 out of 5 stars5/5 (3)
- Circulation or Peripheral CirculationDocument3 pagesCirculation or Peripheral CirculationOldy MutiaraNo ratings yet
- CirculationDocument3 pagesCirculationAIKA BAYANo ratings yet
- Lecture 14-Oct.23 Haemodynamics of Circulation, Circulatory Mechanics, ResistanceDocument29 pagesLecture 14-Oct.23 Haemodynamics of Circulation, Circulatory Mechanics, ResistanceAparmitNo ratings yet
- CVS - Circuitery & HemodynamicsDocument44 pagesCVS - Circuitery & Hemodynamicsspitzmark2030No ratings yet
- The Circulatory SystemDocument51 pagesThe Circulatory SystemMohammed ahmedNo ratings yet
- 40.neural (Extrinsic) Regulation of Cardiac Performance, Sympathetci and Parasympathetic Effects. Humoral Factors Affecting Cardiac PerformanceDocument3 pages40.neural (Extrinsic) Regulation of Cardiac Performance, Sympathetci and Parasympathetic Effects. Humoral Factors Affecting Cardiac PerformanceNektarios TsakalosNo ratings yet
- The Circulatory SystemDocument10 pagesThe Circulatory SystemRezki RasyakNo ratings yet
- Peripheral Circulatory CollapseDocument6 pagesPeripheral Circulatory Collapsehurainsahar21No ratings yet
- Lecture-11 The Heart and Cardiovascular SystemDocument9 pagesLecture-11 The Heart and Cardiovascular Systemmdhawan1be20No ratings yet
- Cardiac PhysiologyDocument102 pagesCardiac PhysiologyPeter Larsen100% (1)
- Circulation J Pressure J Flow and Resistance of by YasirDocument47 pagesCirculation J Pressure J Flow and Resistance of by Yasiryasir ishaqNo ratings yet
- Renal TubuleDocument7 pagesRenal Tubulehurainsahar21No ratings yet
- 23circulation Part 1Document13 pages23circulation Part 1Jaydave PatelNo ratings yet
- Physiology of Circulation SystemDocument52 pagesPhysiology of Circulation SystemReni TeeWeeNo ratings yet
- Special Regional CirculationDocument28 pagesSpecial Regional CirculationBenjamin SharonNo ratings yet
- CardioDocument5 pagesCardioMkhize KhabazelaNo ratings yet
- The Cardiovascular System: Blood Vessels: I. Overview of Blood Vessel Structure and FunctionDocument7 pagesThe Cardiovascular System: Blood Vessels: I. Overview of Blood Vessel Structure and FunctionAdam ButterworthNo ratings yet
- The Cardiovascular System - Blood VesselsDocument39 pagesThe Cardiovascular System - Blood VesselsMohamed YousufNo ratings yet
- OpenStax Rice University 6100 Main Street MS-375 Houston, Texas 77005Document31 pagesOpenStax Rice University 6100 Main Street MS-375 Houston, Texas 77005D VNo ratings yet
- Transport in Man NotesDocument9 pagesTransport in Man NotesYeo JhNo ratings yet
- Science of Living Systems: Bio-Thermal-Fluid Sciences: Lecture by Prof. Soumen DasDocument36 pagesScience of Living Systems: Bio-Thermal-Fluid Sciences: Lecture by Prof. Soumen Dashimanshu_agraNo ratings yet
- CV PathoDocument32 pagesCV PathoSaif AliNo ratings yet
- Pathophysiology of Cardiovascular System Arterial Hypertension Heart ArrhythmiasDocument38 pagesPathophysiology of Cardiovascular System Arterial Hypertension Heart ArrhythmiasLunguVictoriaNo ratings yet
- MF-local Circulation 2016Document61 pagesMF-local Circulation 2016OnSolomonNo ratings yet
- Circulatory System of MammalsDocument10 pagesCirculatory System of Mammalsben-jNo ratings yet
- Resp CH 2Document68 pagesResp CH 2Ahmed khanNo ratings yet
- Normal Changes With Aging in The Cardiovascular SystemDocument4 pagesNormal Changes With Aging in The Cardiovascular SystemSheenarieNo ratings yet
- The Cardiovascular System & Physiology of HeartDocument41 pagesThe Cardiovascular System & Physiology of HeartTheresya Pratiwhy KoniyoNo ratings yet
- Circulation and Cardiovascular SystemDocument12 pagesCirculation and Cardiovascular SystemSatria MandalaNo ratings yet
- Blood Pressure HomeostasisDocument7 pagesBlood Pressure HomeostasisHassan Al SinanNo ratings yet
- Blood Circulatory SystemDocument8 pagesBlood Circulatory SystemstephanietoppoNo ratings yet
- Linkage CrossingDocument8 pagesLinkage Crossinghurainsahar21No ratings yet
- Cardiovascular System PDFDocument12 pagesCardiovascular System PDFVon Valentine MhuteNo ratings yet
- Transport SystemDocument31 pagesTransport Systemhurainsahar21No ratings yet
- Cardiovascular System: Iii. Anatomy and PhysiologyDocument7 pagesCardiovascular System: Iii. Anatomy and PhysiologyYam Bite-Size FabelloNo ratings yet
- II. Blood Vessels Physiology By: Dr. Abdulrahman Aqra MD MSCDocument12 pagesII. Blood Vessels Physiology By: Dr. Abdulrahman Aqra MD MSCapi-290338101No ratings yet
- Coronary CirculationDocument4 pagesCoronary CirculationReena MathewNo ratings yet
- 565656Document61 pages565656Bashar AntriNo ratings yet
- Cardiovascular System - Lecture IIDocument5 pagesCardiovascular System - Lecture IIpragantraNo ratings yet
- Cardiovascular System: Presented by DR Aparna Ramachandran Mds 1 Dept of Public Health DentistryDocument73 pagesCardiovascular System: Presented by DR Aparna Ramachandran Mds 1 Dept of Public Health DentistryAparna RamachandranNo ratings yet
- Cardio Vascular SystemDocument8 pagesCardio Vascular SystemguptaasitNo ratings yet
- Cardiac CycleDocument3 pagesCardiac CycleKhadijah HabeebahNo ratings yet
- Cardiovascular System Lecture 1 2020 2021Document5 pagesCardiovascular System Lecture 1 2020 2021olawandeilo123No ratings yet
- Cardiovascular and Circulatory SystemDocument29 pagesCardiovascular and Circulatory SystemfayeleechaiyapornkulNo ratings yet
- The Cardiovascular System & Physiology of Heart: Presented By: Bhupendra Kumar Integrated M.TechDocument41 pagesThe Cardiovascular System & Physiology of Heart: Presented By: Bhupendra Kumar Integrated M.TechVignesh AshokNo ratings yet
- Vascular Distensibility and Functions of The Arterial and 1Document83 pagesVascular Distensibility and Functions of The Arterial and 1gabby chaanNo ratings yet
- Pulmonary CirculationDocument4 pagesPulmonary CirculationDr Md Abedur RahmanNo ratings yet
- Biology - Circulatory System Notes 1Document13 pagesBiology - Circulatory System Notes 1archit.kulkarni7756No ratings yet
- 3 Circulation in ManDocument7 pages3 Circulation in ManAnasatcia Mcpherson 9c FNo ratings yet
- Unit Seven: Cardiovascular SystemDocument10 pagesUnit Seven: Cardiovascular SystemKath RubioNo ratings yet
- Cardiac Control CentreDocument8 pagesCardiac Control CentregeorgeloxdaleNo ratings yet
- KP 1.3.2.6 Prinsip Hemodinamik 1 (2 Jam)Document49 pagesKP 1.3.2.6 Prinsip Hemodinamik 1 (2 Jam)Try MutiaraNo ratings yet
- (Lecture 7) The Cardiovascular, Respiratory and Lymphatic SystemDocument32 pages(Lecture 7) The Cardiovascular, Respiratory and Lymphatic SystemKasraSrNo ratings yet
- Anatomy McqsDocument3 pagesAnatomy McqsMunazzaNo ratings yet
- 3 Cardiovascular PhysiologyDocument79 pages3 Cardiovascular PhysiologyJose Luna100% (3)
- The Abdomen 98 QDocument78 pagesThe Abdomen 98 QNahidNo ratings yet
- Lymphatic System OutlineDocument4 pagesLymphatic System OutlineMelljonhNo ratings yet
- THE Lymphatic System & Body DefensesDocument39 pagesTHE Lymphatic System & Body DefensesSaiful AmriNo ratings yet
- Schwartz 9th Ed: Chapter Outline: The SpleenDocument22 pagesSchwartz 9th Ed: Chapter Outline: The SpleenKathryn Reunilla100% (1)
- TO Splenectomy 2Document31 pagesTO Splenectomy 2sphericalfaNo ratings yet
- Management of Splenic Injury in The Adult Trauma PatientDocument17 pagesManagement of Splenic Injury in The Adult Trauma PatientArnaldo Santizo SáenzNo ratings yet
- Reflexology Association of Canada - Foot Chart: Plantar ViewDocument1 pageReflexology Association of Canada - Foot Chart: Plantar ViewprabhaNo ratings yet
- Radiology Presentation FinalDocument45 pagesRadiology Presentation FinalKhupe KafundaNo ratings yet
- Aetiology Pathology and Treatment of Blood Stasis PDFDocument248 pagesAetiology Pathology and Treatment of Blood Stasis PDFMiguel Sabaj100% (1)
- Five ElementsDocument21 pagesFive ElementsAry Nogueira100% (2)
- Nigella SativaDocument9 pagesNigella SativaoshinNo ratings yet
- Physiology CH 15 - Vascular Distensibility and FunctionDocument31 pagesPhysiology CH 15 - Vascular Distensibility and FunctionDaniel AdamsNo ratings yet
- Classic Autopsy TechniquesDocument27 pagesClassic Autopsy TechniquesYenny BelindaNo ratings yet
- NICE 2018 Book of Abstracts PDFDocument408 pagesNICE 2018 Book of Abstracts PDFMaria Teresa PulidoNo ratings yet
- Fast Protocol PDFDocument103 pagesFast Protocol PDFSimina SerbanNo ratings yet
- Astral BodiesDocument52 pagesAstral BodiesHaris LiviuNo ratings yet
- E L e N T ADocument34 pagesE L e N T ANika MtsituriNo ratings yet
- PathoLOGY MCQDocument67 pagesPathoLOGY MCQHUSAMNo ratings yet
- Abdominal Visceral Pain: Clinical AspectDocument4 pagesAbdominal Visceral Pain: Clinical AspectMagdy GabrNo ratings yet
- Blunt Abdominal TraumaDocument65 pagesBlunt Abdominal TraumaAmit MishraNo ratings yet
- 2.histologi DarahDocument38 pages2.histologi DarahIrfanNo ratings yet
- Splenomegaly 180319023541Document21 pagesSplenomegaly 180319023541Mohd ShoaibNo ratings yet
- Anatomy of StomachDocument4 pagesAnatomy of StomachIntan Putri Maisarah100% (1)
- Adjectives PDFDocument2 pagesAdjectives PDFPrescQuiNo ratings yet
- The Anatomy and Histology of The Spleen. Differentiation of The Ectoderm PDFDocument5 pagesThe Anatomy and Histology of The Spleen. Differentiation of The Ectoderm PDFredderdatNo ratings yet
- Anatomy Liver, Biliary, Pancreas and Spleen 2022Document58 pagesAnatomy Liver, Biliary, Pancreas and Spleen 2022Osama AbdelazizNo ratings yet
- CANINE-Management Protocol For Acute Gastric Dilatation-Volvulus Syndrom in DogsDocument7 pagesCANINE-Management Protocol For Acute Gastric Dilatation-Volvulus Syndrom in Dogstaner_soysurenNo ratings yet