Download as pptx, pdf, or txt
You might also like
- Bio Factsheet Chloroplasts - Structure and Function Number 198 WWW - Curriculum-Press - Co.ukDocument4 pagesBio Factsheet Chloroplasts - Structure and Function Number 198 WWW - Curriculum-Press - Co.ukbekoNo ratings yet
- Immune Response To VirusDocument25 pagesImmune Response To VirusWindy Hapsari50% (2)
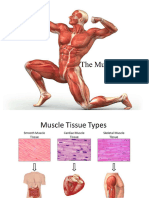
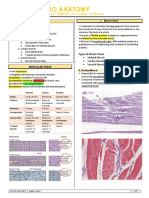
- MuscleDocument11 pagesMuscleJeff ParkNo ratings yet
- B Muscle Tissue HistologyDocument55 pagesB Muscle Tissue HistologyAffie SaikolNo ratings yet
- Skeletal Muscle StructureDocument38 pagesSkeletal Muscle Structuretoyy100% (1)
- Muscular TDocument18 pagesMuscular TSumayya KabeerNo ratings yet
- Wa0038Document10 pagesWa0038Shehzad QureshiNo ratings yet
- 589bc774 Df1d 47c6 Bc97 B5fd73030ca6muscle PhysiologyDocument3 pages589bc774 Df1d 47c6 Bc97 B5fd73030ca6muscle PhysiologyMalti SharmaNo ratings yet
- Muscle PhysiologyDocument24 pagesMuscle PhysiologyArmando AlehandroNo ratings yet
- 6.muscular TissueDocument40 pages6.muscular Tissuemero wowNo ratings yet
- Skeletal MuscleDocument98 pagesSkeletal MuscleMoon Moon100% (1)
- Histology of Cardiac and Smooth MuscleDocument73 pagesHistology of Cardiac and Smooth MuscleShehryar AbbasNo ratings yet
- Histology of Cardiac and Smooth MuscleDocument73 pagesHistology of Cardiac and Smooth MuscleShehryar AbbasNo ratings yet
- Muscle Tissue-Wl06.11Document54 pagesMuscle Tissue-Wl06.11api-19641337No ratings yet
- Microscopic Anatomy of A Skeletal Muscle FiberDocument33 pagesMicroscopic Anatomy of A Skeletal Muscle FiberKumar MSNo ratings yet
- Histology of MuscleDocument49 pagesHistology of MuscleTewachew Dessie100% (1)
- Histo MuscleDocument11 pagesHisto MuscleaikaNo ratings yet
- Muscle Intro-Skeletal Muscle IDocument31 pagesMuscle Intro-Skeletal Muscle IPehovelo JohannesNo ratings yet
- Muscle DentalDocument74 pagesMuscle Dentalapi-3710331No ratings yet
- Muscular SystemDocument10 pagesMuscular SystemMarie Veatrice JacomilleNo ratings yet
- Physiology of Muscle For AnaestDocument96 pagesPhysiology of Muscle For AnaestBahredin AbdellaNo ratings yet
- 04muscle TissueDocument39 pages04muscle Tissueapi-19916399No ratings yet
- ANPH Midterm NotesDocument44 pagesANPH Midterm NotesClaudine Jo B. TalabocNo ratings yet
- Muscle Tissue: - Skeletal Muscle - Cardiac Muscle - Smooth MuscleDocument36 pagesMuscle Tissue: - Skeletal Muscle - Cardiac Muscle - Smooth MuscleDentist Dina Samy100% (1)
- Functional Anatomy - ContentDocument14 pagesFunctional Anatomy - ContentxanaduNo ratings yet
- Chapter 10Document5 pagesChapter 10Nona PeriarceNo ratings yet
- Contraction of Skeletal Muscle (1) : Narrated LectureDocument45 pagesContraction of Skeletal Muscle (1) : Narrated LectureDeanNo ratings yet
- Chapter 10: Muscle Tissue: VoluntaryDocument13 pagesChapter 10: Muscle Tissue: VoluntaryClyde BaltazarNo ratings yet
- Muscular TissuesDocument46 pagesMuscular TissuesDAVE CANALETANo ratings yet
- Lecture 8 (Muscular System Lecture 2)Document43 pagesLecture 8 (Muscular System Lecture 2)Husnain WattoNo ratings yet
- Ucpphysio 313 APharm Dfinal F18Document8 pagesUcpphysio 313 APharm Dfinal F18Huzaifa ChauhanNo ratings yet
- Muscle Pgy - 222Document48 pagesMuscle Pgy - 222SAVIOUR BANDANo ratings yet
- Chapter 7 - Muscular SystemDocument29 pagesChapter 7 - Muscular SystemlNo ratings yet
- Célula Muscular, Macro y MicrobiologicoDocument11 pagesCélula Muscular, Macro y MicrobiologicoimanolNo ratings yet
- Muscle Tissues: Mrs. Ofelia Solano SaludarDocument65 pagesMuscle Tissues: Mrs. Ofelia Solano SaludarYulius SamuelNo ratings yet
- Muscle Lecture On (Excitable Tissue)Document126 pagesMuscle Lecture On (Excitable Tissue)Muhammed ZainabNo ratings yet
- LPT 321Document103 pagesLPT 321rahul roshanNo ratings yet
- Muscle Tissue LectureDocument34 pagesMuscle Tissue LectureurjaNo ratings yet
- Muscles TransesDocument3 pagesMuscles TranseskimeehhNo ratings yet
- Muscle PhysiologyDocument88 pagesMuscle PhysiologyAftab MughalNo ratings yet
- The Muscular System: DR Faisal Institute of Physiology and PharmacologyDocument61 pagesThe Muscular System: DR Faisal Institute of Physiology and PharmacologyTask BirdNo ratings yet
- Lec4 Muscles CompressedDocument27 pagesLec4 Muscles Compressedmohammedbaker1q2w3eNo ratings yet
- Muscle PhysiologyDocument34 pagesMuscle Physiologymadeha goharNo ratings yet
- 6 - Muscle Tissue DR - Vera 2021-2Document45 pages6 - Muscle Tissue DR - Vera 2021-2Wilmarilys Ramos MartellNo ratings yet
- Muscular TissueDocument65 pagesMuscular TissueMitzel SapaloNo ratings yet
- Contractile CellsDocument116 pagesContractile Cellsnav_malhiNo ratings yet
- Revision Notes On Locomotion and Movement For NEET 2024 - Free PDF DownloadDocument11 pagesRevision Notes On Locomotion and Movement For NEET 2024 - Free PDF DownloadHaider KaziNo ratings yet
- ST - of Myofilaments..Document10 pagesST - of Myofilaments..Mohammed HamadaNo ratings yet
- Chapter 5 Excitable Tissue Muscle (Ganong) QuianoDocument5 pagesChapter 5 Excitable Tissue Muscle (Ganong) Quianoim. EliasNo ratings yet
- Muscular SystemDocument10 pagesMuscular Systematerbat07No ratings yet
- 20 Locomotion N Movement PPT - Part 2Document14 pages20 Locomotion N Movement PPT - Part 2sumeeshkNo ratings yet
- HAPP Week VII XDocument32 pagesHAPP Week VII Xxuxi dulNo ratings yet
- Nerve and Muscle Lecture 6 paraDocument24 pagesNerve and Muscle Lecture 6 paraTatenda SibandaNo ratings yet
- Histo 1.9 Muscle TissueDocument7 pagesHisto 1.9 Muscle TissueMonesa Christy VillanuevaNo ratings yet
- Unit 3-Part 1-Skeletal Muscle Anatomy, Sarcomeres, and EC Coupling-SDocument32 pagesUnit 3-Part 1-Skeletal Muscle Anatomy, Sarcomeres, and EC Coupling-Sgdhanoa55No ratings yet
- MascularDocument39 pagesMascularAive BelistaNo ratings yet
- Anaphy-Muscular HandoutDocument7 pagesAnaphy-Muscular HandoutAEGEL DARO TORRALBANo ratings yet
- Muscle Physiology: Human Physiology By-Liul M. ForDocument63 pagesMuscle Physiology: Human Physiology By-Liul M. ForKhant Si ThuNo ratings yet
- NotesDocument15 pagesNotesGizem OsmanogluNo ratings yet
- Contraction of Skeletal MuscleDocument64 pagesContraction of Skeletal MuscleFhrecy Ann ConsularNo ratings yet
- Evolution of cAMP Dependent Protein Kinase PKA CATALYTIC UNIT ISOFORMS Pone.0181091Document17 pagesEvolution of cAMP Dependent Protein Kinase PKA CATALYTIC UNIT ISOFORMS Pone.0181091Sudhir RavindranNo ratings yet
- Cellular Respiration 9-12Document56 pagesCellular Respiration 9-12David BayaniNo ratings yet
- Cytokine Protocols - MArc LeyDocument247 pagesCytokine Protocols - MArc LeyCruz-Clara100% (1)
- Preparation of Stained Temporary Mounts of Onion PeelDocument2 pagesPreparation of Stained Temporary Mounts of Onion PeelAnirudh100% (1)
- GlycolysisDocument7 pagesGlycolysisTahirat NasiruNo ratings yet
- Biology Unitwise MCQ - S (MCAT)Document41 pagesBiology Unitwise MCQ - S (MCAT)M Noaman Akbar100% (1)
- Drug Discovery: Targeting Inflammation in DiabetesDocument1 pageDrug Discovery: Targeting Inflammation in Diabeteschanpreet12No ratings yet
- Metabolism of Ketone BodiesDocument22 pagesMetabolism of Ketone BodiesUbaid AhmedNo ratings yet
- Human Histology: Connective Tissue WorksheetDocument4 pagesHuman Histology: Connective Tissue WorksheetNoriel LacsinaNo ratings yet
- M.Prasad Naidu: MSC Medical Biochemistry, Ph.D.Research ScholarDocument39 pagesM.Prasad Naidu: MSC Medical Biochemistry, Ph.D.Research ScholarRiska Resty WasitaNo ratings yet
- B. Exon: C. Transcription, TranslationDocument5 pagesB. Exon: C. Transcription, TranslationRuhna BaigNo ratings yet
- Erythrocyte Production and DestructionDocument5 pagesErythrocyte Production and DestructionJoanne JardinNo ratings yet
- X Bio Masterkey Differences 22 - 234Document40 pagesX Bio Masterkey Differences 22 - 234Varenayam editzNo ratings yet
- Endomembrane SystemDocument26 pagesEndomembrane SystemPreeti SainiNo ratings yet
- Unit Vi: Storage and Expression of Genetic Information DNA Structure, Replication, and Repair Qingsong WangDocument30 pagesUnit Vi: Storage and Expression of Genetic Information DNA Structure, Replication, and Repair Qingsong WangasaadsarfrazNo ratings yet
- Cell Structure2 FDocument10 pagesCell Structure2 FFred H HalderNo ratings yet
- Selina Concise Biology Solutions Class 6 Chapter 3 Cell - The Structure and FunctionsDocument10 pagesSelina Concise Biology Solutions Class 6 Chapter 3 Cell - The Structure and FunctionsFLANTAMOCKNo ratings yet
- Cellular RespirationDocument27 pagesCellular RespirationAndile SopengNo ratings yet
- Cells and Tissues: Part BDocument25 pagesCells and Tissues: Part BKeanna ZurriagaNo ratings yet
- Bioenergetics and Metabolism - MitochondrionDocument5 pagesBioenergetics and Metabolism - Mitochondrionpaul catalinNo ratings yet
- Cancer Cell and Cell CycleDocument10 pagesCancer Cell and Cell CycleRouie AzucenaNo ratings yet
- Cell DivisionDocument52 pagesCell DivisionRamachandran Ram100% (1)
- Question Paper Unit 2 (6BI02) June 2014Document28 pagesQuestion Paper Unit 2 (6BI02) June 2014Natalie RossetteNo ratings yet
- Siklus KalvinDocument5 pagesSiklus Kalvinnovani florensiaNo ratings yet
- 2.1 Cell Structure and FunctionDocument39 pages2.1 Cell Structure and Functionforyourhonour wongNo ratings yet
- BY: Sunita F1C1 16 064: Chemistry Department Mathematics and Natural Sciences Faculty Halu Oleo University Kendari 2018Document26 pagesBY: Sunita F1C1 16 064: Chemistry Department Mathematics and Natural Sciences Faculty Halu Oleo University Kendari 2018Azka Zaina ArdiningrumNo ratings yet
- Name: Nur Haura Zhafirah Lubis Olinda Calista Yetri ClassDocument8 pagesName: Nur Haura Zhafirah Lubis Olinda Calista Yetri ClassNur HauraNo ratings yet
- MD3 Immunology Final Exam ReviewDocument28 pagesMD3 Immunology Final Exam Reviewdavid100% (1)