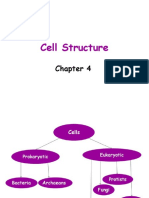
Download as pptx, pdf, or txt
You might also like
- 4 Corners Gold Rush Lesson PlanDocument3 pages4 Corners Gold Rush Lesson Planapi-267003013No ratings yet
- Fundamentals of Nursing PDFDocument18 pagesFundamentals of Nursing PDFJamaica Manuel Iglesias100% (6)
- Holistic Self Health CareDocument80 pagesHolistic Self Health Careadkittipong100% (1)
- Unit 15 Cell Junction DhanDocument18 pagesUnit 15 Cell Junction DhanSarah PavuNo ratings yet
- 19) Integrating Cells Into TissuesDocument37 pages19) Integrating Cells Into TissuesRNo ratings yet
- Cell JunctionsDocument24 pagesCell JunctionsNajlaNo ratings yet
- Cell JunctionDocument63 pagesCell JunctionsphuyalNo ratings yet
- Cell-to-Cell and Cell-to-Matrix Adhesions: Fibronectin?Document46 pagesCell-to-Cell and Cell-to-Matrix Adhesions: Fibronectin?MinarsihNo ratings yet
- BBS1 HS K4 Cell Junction Intercellular CommunicationDocument26 pagesBBS1 HS K4 Cell Junction Intercellular CommunicationVania IlmiNo ratings yet
- Physiological Processes Where Cell Adhesion Is ImportantDocument42 pagesPhysiological Processes Where Cell Adhesion Is ImportantJT92No ratings yet
- Cell-Cell Junctions: Bhavtosh A. Kikani, PHD, Icar-NetDocument16 pagesCell-Cell Junctions: Bhavtosh A. Kikani, PHD, Icar-NetB KikaniNo ratings yet
- of Cell Adhesion and Cell JunctionsDocument50 pagesof Cell Adhesion and Cell Junctionsayad ghawiNo ratings yet
- L4 - Cell JunctionDocument15 pagesL4 - Cell Junctionطارق ابولبدةNo ratings yet
- Cytoskeleton, Junctions, Fibroblast and ECMDocument38 pagesCytoskeleton, Junctions, Fibroblast and ECMManoj HumagainNo ratings yet
- CelladhesionDocument51 pagesCelladhesionNina Difla MuflikhahNo ratings yet
- Celladhesion Epithel KissADocument51 pagesCelladhesion Epithel KissAamdusuf33No ratings yet
- CellDocument47 pagesCellAnami NizamNo ratings yet
- Introduction To Histology and Epithelium TissueDocument48 pagesIntroduction To Histology and Epithelium TissuejabirNo ratings yet
- Cell Bio 3 Cell Junctions 2021 PDFDocument38 pagesCell Bio 3 Cell Junctions 2021 PDFaboodmr8nblahNo ratings yet
- Cell JunctionsDocument40 pagesCell JunctionsAhmad MursyidNo ratings yet
- Geral Aula #4a - Bioquí Mica e Biologia CelularDocument40 pagesGeral Aula #4a - Bioquí Mica e Biologia CelularsusanajpNo ratings yet
- Cell Adhesion Molecules 300107Document32 pagesCell Adhesion Molecules 300107V Lee 'Nozhat'100% (1)
- Cytoskeleton 1Document75 pagesCytoskeleton 1Abdul Rehman IrfanNo ratings yet
- Junctions - 2008 ArisDocument34 pagesJunctions - 2008 ArisAmardeep SinghNo ratings yet
- Cell JunctionsDocument56 pagesCell JunctionsDrGagandeep Singh100% (3)
- Cell Junction, Adhesion and Extracellular MatrixDocument14 pagesCell Junction, Adhesion and Extracellular Matrixseenu mohapatraNo ratings yet
- LS7C Week 1B Pre-Class Reading GuideDocument3 pagesLS7C Week 1B Pre-Class Reading GuidekaedechungNo ratings yet
- Cell Junctions and Cell AdhesionDocument31 pagesCell Junctions and Cell AdhesionCitrawati Dyah Kencono WunguNo ratings yet
- Lecture 2-Cell Structure - LZDocument44 pagesLecture 2-Cell Structure - LZyusm2312No ratings yet
- The CytoskeletonDocument63 pagesThe CytoskeletonAmara TargaryenNo ratings yet
- BM 1 1 CytoskeletonDocument24 pagesBM 1 1 CytoskeletonSanthoshi Sadhanaa SankarNo ratings yet
- Cell Surface receptorsMCBA P2 TDocument16 pagesCell Surface receptorsMCBA P2 TMatende husseinNo ratings yet
- 05lect Adhesion Summary2017Document5 pages05lect Adhesion Summary2017Dan ChoiNo ratings yet
- Cell Junctions: Cell Junctions: Structure & Role: Dr. R. Debnath Assoc. Professor Deptt. of Zoology M.B.B. CollegeDocument74 pagesCell Junctions: Cell Junctions: Structure & Role: Dr. R. Debnath Assoc. Professor Deptt. of Zoology M.B.B. Collegefmukuka12No ratings yet
- Cytoskeleton Motor Proteins and ECM - SCDocument77 pagesCytoskeleton Motor Proteins and ECM - SCAlice RizzoNo ratings yet
- Cells AdhesionDocument18 pagesCells AdhesionAvesta AbbasNo ratings yet
- Cell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemDocument70 pagesCell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemGovinda BiswasNo ratings yet
- Adhesion LectureDocument26 pagesAdhesion Lecturedoubleyouem2003No ratings yet
- Cell JunctionDocument10 pagesCell JunctiongsvssumaNo ratings yet
- Lecture 1 Cell and Molecular Biology 1Document38 pagesLecture 1 Cell and Molecular Biology 1Martin HangasNo ratings yet
- Cell Junction (Intercellular Bridges)Document23 pagesCell Junction (Intercellular Bridges)Aishwarya MohanNo ratings yet
- Activity No. 1-Cell ComponentsDocument3 pagesActivity No. 1-Cell ComponentsKyla QuilantangNo ratings yet
- Class 19 CellBiology MB 2023 2024Document20 pagesClass 19 CellBiology MB 2023 2024André MatosNo ratings yet
- Cell JunctionsDocument30 pagesCell JunctionsNavodit GoelNo ratings yet
- Intercellular JunctionsDocument20 pagesIntercellular Junctionsjohnthar929No ratings yet
- Partoza, J WS Cel - MolDocument6 pagesPartoza, J WS Cel - MolJenny PartozaNo ratings yet
- Cell JunctionsDocument34 pagesCell JunctionsDrAmit Gaba Mds100% (1)
- 03 Epithelium 2022Document78 pages03 Epithelium 2022AlkadafeNo ratings yet
- Intercellular JunctionsDocument5 pagesIntercellular Junctionspetricia johnsonNo ratings yet
- Cy To SkeletonDocument21 pagesCy To SkeletonJisna JaisonNo ratings yet
- Biomolecules & Cells Biol10002 Lectures 1-9 Cell Biology Geoff Mcfadden, RM 211 Biosciences 2 (Old Botany)Document41 pagesBiomolecules & Cells Biol10002 Lectures 1-9 Cell Biology Geoff Mcfadden, RM 211 Biosciences 2 (Old Botany)waimoeNo ratings yet
- A Cell Is The Smallest Unit of A Living ThingDocument4 pagesA Cell Is The Smallest Unit of A Living ThingJONAVIE DEMALATANo ratings yet
- Cell Cytoskeleton, Cell JunctionsDocument46 pagesCell Cytoskeleton, Cell JunctionsAhmed khanNo ratings yet
- Chapter 4 BIO 1510Document30 pagesChapter 4 BIO 1510Chachi CNo ratings yet
- Cell CommunicationDocument27 pagesCell Communicationapi-292966101No ratings yet
- Integrasi Sel Dalam Jaringan (Kuliah) - 2 - 2Document125 pagesIntegrasi Sel Dalam Jaringan (Kuliah) - 2 - 2komang nickoNo ratings yet
- Cancer CellsDocument2 pagesCancer CellsShubhendu Kumar TripathiNo ratings yet
- In Molecular Cell Biology: January 1995Document39 pagesIn Molecular Cell Biology: January 1995avinmanzanoNo ratings yet
- Bio2000 Notes-2004-2Document21 pagesBio2000 Notes-2004-2api-3700537No ratings yet
- CADHERINS FOR CELL ADHESION batulanDocument5 pagesCADHERINS FOR CELL ADHESION batulanBonna Jay GuinsodNo ratings yet
- 2 Basal Lateral Apical Modifications and Connective TissueDocument84 pages2 Basal Lateral Apical Modifications and Connective TissueDeniz asmazNo ratings yet
- Chapter 2 - NotesDocument22 pagesChapter 2 - NotesanusoumyaNo ratings yet
- RM Aiats 1 2022Document24 pagesRM Aiats 1 2022Anyone Can CookNo ratings yet
- 206 1 Summer Internship Programme 2020Document1 page206 1 Summer Internship Programme 2020Anyone Can CookNo ratings yet
- Ayushi VakilDocument16 pagesAyushi VakilAnyone Can CookNo ratings yet
- Swarnali DasguptaDocument16 pagesSwarnali DasguptaAnyone Can CookNo ratings yet
- 218 - 1 - Summer Internship Programme 2021Document3 pages218 - 1 - Summer Internship Programme 2021Anyone Can CookNo ratings yet
- Manasa BalakrishnanDocument14 pagesManasa BalakrishnanAnyone Can CookNo ratings yet
- Bishwarup SarkarDocument18 pagesBishwarup SarkarAnyone Can CookNo ratings yet
- Sandip BambhaniyaDocument13 pagesSandip BambhaniyaAnyone Can CookNo ratings yet
- Ensuring Safety and Cleanliness in The SalonDocument21 pagesEnsuring Safety and Cleanliness in The SalonMa Christina Encinas83% (6)
- Livebook ACBDocument420 pagesLivebook ACBShubhangi JagtapNo ratings yet
- Background of The StudyDocument13 pagesBackground of The Studyjeffrey rodriguezNo ratings yet
- Seed Extraction MethodsDocument3 pagesSeed Extraction MethodsPreetam NayakNo ratings yet
- Sdfsdynamic AuscultationDocument3 pagesSdfsdynamic AuscultationrichardNo ratings yet
- TN-4620190912122 Form21B Signed PDFDocument1 pageTN-4620190912122 Form21B Signed PDFDHEEN MEDICALSNo ratings yet
- IFU Acticor 439128-F EsDocument143 pagesIFU Acticor 439128-F EsMartin ArrietaNo ratings yet
- PROJECT On Tea LeavesDocument15 pagesPROJECT On Tea LeavesBharat Singh83% (6)
- 793-P-1C Relay Data SheetDocument6 pages793-P-1C Relay Data SheetsendmebooksNo ratings yet
- Electrical Commissioning Checklist Non LeedDocument7 pagesElectrical Commissioning Checklist Non Leedcaleb isukuruNo ratings yet
- Pentingnya Memelihara Kebersihan Dan Keamanan Lingkungan Secarapartisipatif Demi Meningkatkan Gotong Royong Dan Kualitas Hidup WargaDocument6 pagesPentingnya Memelihara Kebersihan Dan Keamanan Lingkungan Secarapartisipatif Demi Meningkatkan Gotong Royong Dan Kualitas Hidup WargaMey MeyNo ratings yet
- Bence Bays Resume July 2015Document2 pagesBence Bays Resume July 2015api-292242662No ratings yet
- Dietary Computation For Pregnant ClientDocument12 pagesDietary Computation For Pregnant ClientLuis WashingtonNo ratings yet
- Crew Habitability On Mobile Offshore Drilling Units (Modus) : Guide ForDocument87 pagesCrew Habitability On Mobile Offshore Drilling Units (Modus) : Guide ForReza BabriNo ratings yet
- PHAS0027 RevisionDocument21 pagesPHAS0027 Revisionunknown.unknown9901No ratings yet
- Material Sub Group Item CodeDocument2,818 pagesMaterial Sub Group Item Codegouri gouriNo ratings yet
- 2442 4348 2 PBDocument8 pages2442 4348 2 PBdev iNo ratings yet
- Frenkel's ExerciseDocument40 pagesFrenkel's ExerciseManiu EmeseNo ratings yet
- Ethics in HRMDocument7 pagesEthics in HRMVinay RamaneNo ratings yet
- Final LIST OF IMPORTANT GRID ELEMENTS Updated On May 2020 PDFDocument74 pagesFinal LIST OF IMPORTANT GRID ELEMENTS Updated On May 2020 PDFbhargavNo ratings yet
- GRES Integrated Energy Storage SystemDocument33 pagesGRES Integrated Energy Storage SystemVadim PopovichNo ratings yet
- Training - Cga ApplicationDocument34 pagesTraining - Cga ApplicationSubhan Muhammad100% (1)
- Starkville Dispatch Eedition 7-29-20Document16 pagesStarkville Dispatch Eedition 7-29-20The DispatchNo ratings yet
- II 2015 4Document283 pagesII 2015 4tuni santeNo ratings yet
- Chapter4 Questions WithoutSupersonicDocument5 pagesChapter4 Questions WithoutSupersonicsh1999No ratings yet
- CAT MENTAL HEALTH FOR Community Health YEAR 4Document4 pagesCAT MENTAL HEALTH FOR Community Health YEAR 4Dennis ButtoNo ratings yet
- Proforma A1: Residential/Domicile CertificateDocument6 pagesProforma A1: Residential/Domicile CertificateSamim ParvezNo ratings yet