
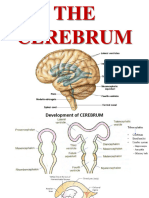
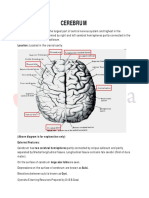
ANATOMY OF CEREBRAL CORTEX
ANATOMY OF CEREBRAL CORTEX
You might also like
- 2002 System Wiring Diagrams Toyota CorollaDocument41 pages2002 System Wiring Diagrams Toyota Corollakabanek75% (4)
- Unlock Your Macro Types - Training PlansDocument21 pagesUnlock Your Macro Types - Training Plansrfzfwn8nf4No ratings yet
- Test Bank For Essentials of Genetics 10th Edition William S KlugDocument9 pagesTest Bank For Essentials of Genetics 10th Edition William S KlugBarbara Hinkle100% (41)
- Zifeng Tower - Ramazan SariDocument2 pagesZifeng Tower - Ramazan SariNadya Pricilia100% (1)
- Neuroanatomy Revision Notes PDFDocument7 pagesNeuroanatomy Revision Notes PDFTanya Tanu100% (2)
- Bells PalsyDocument15 pagesBells PalsyJoseph NicdaoNo ratings yet
- BookofRaziel PDFDocument74 pagesBookofRaziel PDFLoli Darriba Perez100% (1)
- English File Fourth Edition Intermediate WorkbookDocument82 pagesEnglish File Fourth Edition Intermediate WorkbookАнатоль БумберсNo ratings yet
- Cerebrum AnatomyDocument40 pagesCerebrum AnatomyDr. Thangaraj MDNo ratings yet
- Telencephalon - General OrganizationDocument23 pagesTelencephalon - General OrganizationMirko BelanNo ratings yet
- l15 CerebrumDocument31 pagesl15 CerebrumDr. Pinki RaiNo ratings yet
- Cerebrum 2Document67 pagesCerebrum 2Syed Abdul BasitNo ratings yet
- L15 CerebrumDocument27 pagesL15 CerebrumEvilneko1No ratings yet
- NS 5. Cerebrum SEPDocument28 pagesNS 5. Cerebrum SEPsanthiya rajendranNo ratings yet
- 7 Cerebrum 2022Document36 pages7 Cerebrum 2022Gregory WigginsNo ratings yet
- CerebellumDocument28 pagesCerebellumxwrmaliaramNo ratings yet
- A Complete Laboratory Guide To Module 13Document30 pagesA Complete Laboratory Guide To Module 132012000600No ratings yet
- Neuroanatomy for ModularDocument100 pagesNeuroanatomy for Modularusamahh4334No ratings yet
- External Features of Cerebral HemisphereDocument15 pagesExternal Features of Cerebral HemisphereExtra74 ExtraNo ratings yet
- Cerebral Hemispheres 2018 PDFDocument69 pagesCerebral Hemispheres 2018 PDFlailatul husnaNo ratings yet
- CerebrumDocument59 pagesCerebrumKamlesh DuggaNo ratings yet
- II Sem Lecture Pysiotherapy, BS, SS, 8.05.2020Document19 pagesII Sem Lecture Pysiotherapy, BS, SS, 8.05.2020Alexander VonPatrickNo ratings yet
- Cerebral Cortex 1Document37 pagesCerebral Cortex 1Dorin PathakNo ratings yet
- Brain Anatomy HemispheresDocument37 pagesBrain Anatomy HemispheresSasikala MohanNo ratings yet
- 1A Head and Neck IntroductionDocument17 pages1A Head and Neck IntroductionPataraza, Louis Joshua E.No ratings yet
- Neural Basis 5Document24 pagesNeural Basis 5zainabbu2023No ratings yet
- CerebrumDocument51 pagesCerebrumbella astriniNo ratings yet
- Cerebrum +pons LectureDocument53 pagesCerebrum +pons LectureIsrael Olusegun100% (1)
- Temporal Lobe Abhishek - LatestDocument81 pagesTemporal Lobe Abhishek - LatestMadhushree N SwamyNo ratings yet
- Parietal LobeDocument29 pagesParietal LobeAdrishyaNo ratings yet
- Normal Brain AnatomyDocument85 pagesNormal Brain AnatomyYashan SharmaNo ratings yet
- Cerebrum: By: Pedro E. Ampil, M.D.FPSA Department of Human Structural Biology Our Lady of Fatima UniversityDocument29 pagesCerebrum: By: Pedro E. Ampil, M.D.FPSA Department of Human Structural Biology Our Lady of Fatima UniversityPlaza Jeanine LouiseNo ratings yet
- Cranial BaseDocument25 pagesCranial BasesealovesriverNo ratings yet
- CerebrumDocument12 pagesCerebrumDr santoshNo ratings yet
- Lec 10Document35 pagesLec 10Azhar MehmoodNo ratings yet
- The Brain CerebrumDocument18 pagesThe Brain CerebrumMaquirang Cris JaisonNo ratings yet
- Growth and Development of Cranial Base: SeminarDocument34 pagesGrowth and Development of Cranial Base: SeminarMNSNo ratings yet
- Cerebellum and Basal Ganglia: Presented byDocument68 pagesCerebellum and Basal Ganglia: Presented byDivya AggarwalNo ratings yet
- Organization of Nervous System CNSDocument35 pagesOrganization of Nervous System CNSPayal MulaniNo ratings yet
- Alberta Stroke Program Early CT Score - 1Document4 pagesAlberta Stroke Program Early CT Score - 1Ardina Nur PramudhitaNo ratings yet
- Cerebellum Top 1Document31 pagesCerebellum Top 1hassaanjamilNo ratings yet
- Cerebellum Diana BASMADocument23 pagesCerebellum Diana BASMAviorel79No ratings yet
- Facial NerveDocument23 pagesFacial NerveAmit SinghNo ratings yet
- SPINAL CORD and TRACTSDocument38 pagesSPINAL CORD and TRACTSJames BastesNo ratings yet
- NEUROANATOMY Lecture # 02 (CEREBRUM)Document12 pagesNEUROANATOMY Lecture # 02 (CEREBRUM)doc-fahad aftabNo ratings yet
- Anatomy and PhysiologyDocument4 pagesAnatomy and PhysiologyMaquirang Cris JaisonNo ratings yet
- 13 Anatomy of The Nervous SystemDocument95 pages13 Anatomy of The Nervous SystemolivNo ratings yet
- Dr. Eyad M. Hussein: Anatomy of The Cerebellum, Basal Nuclei & DiencephalonDocument68 pagesDr. Eyad M. Hussein: Anatomy of The Cerebellum, Basal Nuclei & Diencephalonأمال داودNo ratings yet
- Surgical Anatomy of The BrainDocument38 pagesSurgical Anatomy of The BrainAdeNo ratings yet
- Cerebrum Sulci and Gyri and Circle of WillisDocument53 pagesCerebrum Sulci and Gyri and Circle of Willisgandhiayu100% (1)
- Cerebellum and HypothalamusDocument20 pagesCerebellum and HypothalamusSaviourNo ratings yet
- Writeup Physio Doc Agor 1Document15 pagesWriteup Physio Doc Agor 1Luis PadillaNo ratings yet
- The CerebrumDocument27 pagesThe Cerebrumapi-19916399No ratings yet
- Chapter 11.Neuroanatomy (1)Document79 pagesChapter 11.Neuroanatomy (1)essammuh799No ratings yet
- Localization in Clinical Neurology: Dr. Elsa GeorgeDocument64 pagesLocalization in Clinical Neurology: Dr. Elsa GeorgeElsa GeorgeNo ratings yet
- Gross Anatomy of The BrainDocument64 pagesGross Anatomy of The BrainueumanaNo ratings yet
- 31_NeuroanatomyDocument13 pages31_Neuroanatomykokachithavala7No ratings yet
- Brain NotesDocument5 pagesBrain Notesbithika pathakNo ratings yet
- The CerebellumDocument5 pagesThe CerebellumAndrew BarryNo ratings yet
- The Cerebrum Is Divided Into Two Major PartsDocument4 pagesThe Cerebrum Is Divided Into Two Major PartslisaNo ratings yet
- CerebrumDocument38 pagesCerebrumAuza Moses IbrahimNo ratings yet
- Cerebrum 130906013333Document12 pagesCerebrum 130906013333RajveerNo ratings yet
- Skullbase 190310153626 PDFDocument107 pagesSkullbase 190310153626 PDFVivek DevendarNo ratings yet
- Nro NotessafdaadDocument199 pagesNro NotessafdaadMunawwar AwaNo ratings yet
- Internal Anatomy of The Temporal LobesDocument67 pagesInternal Anatomy of The Temporal LobesemmakooffNo ratings yet
- A New Order of Fishlike Amphibia From the Pennsylvanian of KansasFrom EverandA New Order of Fishlike Amphibia From the Pennsylvanian of KansasNo ratings yet
- Sallowing DisorderDocument40 pagesSallowing Disorderluxmee bhattaraiNo ratings yet
- Somatosensory Evoked PotentialDocument39 pagesSomatosensory Evoked Potentialluxmee bhattaraiNo ratings yet
- Management of perceptual disordersDocument30 pagesManagement of perceptual disordersluxmee bhattaraiNo ratings yet
- PHYSIOLOGY OF MOTOR CONTROL IN ACTION SYSTEMDocument33 pagesPHYSIOLOGY OF MOTOR CONTROL IN ACTION SYSTEMluxmee bhattaraiNo ratings yet
- Development of postural controlDocument44 pagesDevelopment of postural controlluxmee bhattaraiNo ratings yet
- Our Best-Selling Platform Lift: Aritco 7000Document2 pagesOur Best-Selling Platform Lift: Aritco 7000Lê Văn HoạtNo ratings yet
- Physical Pharmaceutics - 1Document15 pagesPhysical Pharmaceutics - 1Odin GODNo ratings yet
- Cellular AberrationDocument8 pagesCellular AberrationRaymund IdicaNo ratings yet
- Altop Industries Limited: Sri Lakshmi Plastic IndustriesDocument3 pagesAltop Industries Limited: Sri Lakshmi Plastic IndustriesSMART T PlacementsNo ratings yet
- Ethernet/Ip: Technical ManualDocument68 pagesEthernet/Ip: Technical ManualDiana HerreraNo ratings yet
- Wlna9ce - Parts ManualDocument24 pagesWlna9ce - Parts Manualsclaveau20No ratings yet
- Nanazoxid 0Document5 pagesNanazoxid 0hnn9gfqzqxNo ratings yet
- The Story That Must Not Be Told by Kavery NambisanDocument6 pagesThe Story That Must Not Be Told by Kavery Nambisanfreelancer800100% (1)
- Comparative Evaluation of Antibacterial Efficacy.9Document6 pagesComparative Evaluation of Antibacterial Efficacy.9Shivani DubeyNo ratings yet
- Aoralscan 2 User Manual+Document8 pagesAoralscan 2 User Manual+MAHESH KULKARNINo ratings yet
- Vegan Mayonnaise - Simple Vegan Blog PDFDocument2 pagesVegan Mayonnaise - Simple Vegan Blog PDFjkoiluNo ratings yet
- Networks 512 CA Test 2Document54 pagesNetworks 512 CA Test 2Thina Siphesihle ZuluNo ratings yet
- LSS Catalogue enDocument68 pagesLSS Catalogue enArzu AliyevaNo ratings yet
- CH 18Document9 pagesCH 18Julie Nix FrazierNo ratings yet
- Masters of Sex 1x01 - PilotDocument66 pagesMasters of Sex 1x01 - PilotettaworNo ratings yet
- Disciplines and Ideas in The Social SciencesDocument18 pagesDisciplines and Ideas in The Social SciencesArjane Grace SullanoNo ratings yet
- Ketone Bodies: DR Anjali SaxenaDocument3 pagesKetone Bodies: DR Anjali Saxenahirendra patelNo ratings yet
- Btech 1 Sem Programming For Problem Solving kcs101 2021Document1 pageBtech 1 Sem Programming For Problem Solving kcs101 2021rishabhchauhan2266No ratings yet
- A Place You Want To Stay at On Your Next TripDocument4 pagesA Place You Want To Stay at On Your Next TripĐoàn Thanh Chúc (FGW HN)No ratings yet
- L A Level Physics A MS Jun 07 Old g481Document88 pagesL A Level Physics A MS Jun 07 Old g481dasha962No ratings yet
- ASU-C-1010 (Receiving Inspection - Bolts, Nuts & Washers For Structural Connections) 14122016Document7 pagesASU-C-1010 (Receiving Inspection - Bolts, Nuts & Washers For Structural Connections) 14122016Bhavanishankar ShettyNo ratings yet
- Annual Day AnchoringDocument5 pagesAnnual Day AnchoringPooja ChuriwalNo ratings yet
- SCP TRPG Cheatsheet Collection v3 SmallerDocument4 pagesSCP TRPG Cheatsheet Collection v3 SmallerNUNUゲイNo ratings yet
- Middle Ear Tumors - Dr. Shivendra PandeyDocument60 pagesMiddle Ear Tumors - Dr. Shivendra PandeyNupur GargNo ratings yet




























































































Download as pptx, pdf, or txt
You might also like
- 2002 System Wiring Diagrams Toyota CorollaDocument41 pages2002 System Wiring Diagrams Toyota Corollakabanek75% (4)
- Unlock Your Macro Types - Training PlansDocument21 pagesUnlock Your Macro Types - Training Plansrfzfwn8nf4No ratings yet
- Test Bank For Essentials of Genetics 10th Edition William S KlugDocument9 pagesTest Bank For Essentials of Genetics 10th Edition William S KlugBarbara Hinkle100% (41)
- Zifeng Tower - Ramazan SariDocument2 pagesZifeng Tower - Ramazan SariNadya Pricilia100% (1)
- Neuroanatomy Revision Notes PDFDocument7 pagesNeuroanatomy Revision Notes PDFTanya Tanu100% (2)
- Bells PalsyDocument15 pagesBells PalsyJoseph NicdaoNo ratings yet
- BookofRaziel PDFDocument74 pagesBookofRaziel PDFLoli Darriba Perez100% (1)
- English File Fourth Edition Intermediate WorkbookDocument82 pagesEnglish File Fourth Edition Intermediate WorkbookАнатоль БумберсNo ratings yet
- Cerebrum AnatomyDocument40 pagesCerebrum AnatomyDr. Thangaraj MDNo ratings yet
- Telencephalon - General OrganizationDocument23 pagesTelencephalon - General OrganizationMirko BelanNo ratings yet
- l15 CerebrumDocument31 pagesl15 CerebrumDr. Pinki RaiNo ratings yet
- Cerebrum 2Document67 pagesCerebrum 2Syed Abdul BasitNo ratings yet
- L15 CerebrumDocument27 pagesL15 CerebrumEvilneko1No ratings yet
- NS 5. Cerebrum SEPDocument28 pagesNS 5. Cerebrum SEPsanthiya rajendranNo ratings yet
- 7 Cerebrum 2022Document36 pages7 Cerebrum 2022Gregory WigginsNo ratings yet
- CerebellumDocument28 pagesCerebellumxwrmaliaramNo ratings yet
- A Complete Laboratory Guide To Module 13Document30 pagesA Complete Laboratory Guide To Module 132012000600No ratings yet
- Neuroanatomy for ModularDocument100 pagesNeuroanatomy for Modularusamahh4334No ratings yet
- External Features of Cerebral HemisphereDocument15 pagesExternal Features of Cerebral HemisphereExtra74 ExtraNo ratings yet
- Cerebral Hemispheres 2018 PDFDocument69 pagesCerebral Hemispheres 2018 PDFlailatul husnaNo ratings yet
- CerebrumDocument59 pagesCerebrumKamlesh DuggaNo ratings yet
- II Sem Lecture Pysiotherapy, BS, SS, 8.05.2020Document19 pagesII Sem Lecture Pysiotherapy, BS, SS, 8.05.2020Alexander VonPatrickNo ratings yet
- Cerebral Cortex 1Document37 pagesCerebral Cortex 1Dorin PathakNo ratings yet
- Brain Anatomy HemispheresDocument37 pagesBrain Anatomy HemispheresSasikala MohanNo ratings yet
- 1A Head and Neck IntroductionDocument17 pages1A Head and Neck IntroductionPataraza, Louis Joshua E.No ratings yet
- Neural Basis 5Document24 pagesNeural Basis 5zainabbu2023No ratings yet
- CerebrumDocument51 pagesCerebrumbella astriniNo ratings yet
- Cerebrum +pons LectureDocument53 pagesCerebrum +pons LectureIsrael Olusegun100% (1)
- Temporal Lobe Abhishek - LatestDocument81 pagesTemporal Lobe Abhishek - LatestMadhushree N SwamyNo ratings yet
- Parietal LobeDocument29 pagesParietal LobeAdrishyaNo ratings yet
- Normal Brain AnatomyDocument85 pagesNormal Brain AnatomyYashan SharmaNo ratings yet
- Cerebrum: By: Pedro E. Ampil, M.D.FPSA Department of Human Structural Biology Our Lady of Fatima UniversityDocument29 pagesCerebrum: By: Pedro E. Ampil, M.D.FPSA Department of Human Structural Biology Our Lady of Fatima UniversityPlaza Jeanine LouiseNo ratings yet
- Cranial BaseDocument25 pagesCranial BasesealovesriverNo ratings yet
- CerebrumDocument12 pagesCerebrumDr santoshNo ratings yet
- Lec 10Document35 pagesLec 10Azhar MehmoodNo ratings yet
- The Brain CerebrumDocument18 pagesThe Brain CerebrumMaquirang Cris JaisonNo ratings yet
- Growth and Development of Cranial Base: SeminarDocument34 pagesGrowth and Development of Cranial Base: SeminarMNSNo ratings yet
- Cerebellum and Basal Ganglia: Presented byDocument68 pagesCerebellum and Basal Ganglia: Presented byDivya AggarwalNo ratings yet
- Organization of Nervous System CNSDocument35 pagesOrganization of Nervous System CNSPayal MulaniNo ratings yet
- Alberta Stroke Program Early CT Score - 1Document4 pagesAlberta Stroke Program Early CT Score - 1Ardina Nur PramudhitaNo ratings yet
- Cerebellum Top 1Document31 pagesCerebellum Top 1hassaanjamilNo ratings yet
- Cerebellum Diana BASMADocument23 pagesCerebellum Diana BASMAviorel79No ratings yet
- Facial NerveDocument23 pagesFacial NerveAmit SinghNo ratings yet
- SPINAL CORD and TRACTSDocument38 pagesSPINAL CORD and TRACTSJames BastesNo ratings yet
- NEUROANATOMY Lecture # 02 (CEREBRUM)Document12 pagesNEUROANATOMY Lecture # 02 (CEREBRUM)doc-fahad aftabNo ratings yet
- Anatomy and PhysiologyDocument4 pagesAnatomy and PhysiologyMaquirang Cris JaisonNo ratings yet
- 13 Anatomy of The Nervous SystemDocument95 pages13 Anatomy of The Nervous SystemolivNo ratings yet
- Dr. Eyad M. Hussein: Anatomy of The Cerebellum, Basal Nuclei & DiencephalonDocument68 pagesDr. Eyad M. Hussein: Anatomy of The Cerebellum, Basal Nuclei & Diencephalonأمال داودNo ratings yet
- Surgical Anatomy of The BrainDocument38 pagesSurgical Anatomy of The BrainAdeNo ratings yet
- Cerebrum Sulci and Gyri and Circle of WillisDocument53 pagesCerebrum Sulci and Gyri and Circle of Willisgandhiayu100% (1)
- Cerebellum and HypothalamusDocument20 pagesCerebellum and HypothalamusSaviourNo ratings yet
- Writeup Physio Doc Agor 1Document15 pagesWriteup Physio Doc Agor 1Luis PadillaNo ratings yet
- The CerebrumDocument27 pagesThe Cerebrumapi-19916399No ratings yet
- Chapter 11.Neuroanatomy (1)Document79 pagesChapter 11.Neuroanatomy (1)essammuh799No ratings yet
- Localization in Clinical Neurology: Dr. Elsa GeorgeDocument64 pagesLocalization in Clinical Neurology: Dr. Elsa GeorgeElsa GeorgeNo ratings yet
- Gross Anatomy of The BrainDocument64 pagesGross Anatomy of The BrainueumanaNo ratings yet
- 31_NeuroanatomyDocument13 pages31_Neuroanatomykokachithavala7No ratings yet
- Brain NotesDocument5 pagesBrain Notesbithika pathakNo ratings yet
- The CerebellumDocument5 pagesThe CerebellumAndrew BarryNo ratings yet
- The Cerebrum Is Divided Into Two Major PartsDocument4 pagesThe Cerebrum Is Divided Into Two Major PartslisaNo ratings yet
- CerebrumDocument38 pagesCerebrumAuza Moses IbrahimNo ratings yet
- Cerebrum 130906013333Document12 pagesCerebrum 130906013333RajveerNo ratings yet
- Skullbase 190310153626 PDFDocument107 pagesSkullbase 190310153626 PDFVivek DevendarNo ratings yet
- Nro NotessafdaadDocument199 pagesNro NotessafdaadMunawwar AwaNo ratings yet
- Internal Anatomy of The Temporal LobesDocument67 pagesInternal Anatomy of The Temporal LobesemmakooffNo ratings yet
- A New Order of Fishlike Amphibia From the Pennsylvanian of KansasFrom EverandA New Order of Fishlike Amphibia From the Pennsylvanian of KansasNo ratings yet
- Sallowing DisorderDocument40 pagesSallowing Disorderluxmee bhattaraiNo ratings yet
- Somatosensory Evoked PotentialDocument39 pagesSomatosensory Evoked Potentialluxmee bhattaraiNo ratings yet
- Management of perceptual disordersDocument30 pagesManagement of perceptual disordersluxmee bhattaraiNo ratings yet
- PHYSIOLOGY OF MOTOR CONTROL IN ACTION SYSTEMDocument33 pagesPHYSIOLOGY OF MOTOR CONTROL IN ACTION SYSTEMluxmee bhattaraiNo ratings yet
- Development of postural controlDocument44 pagesDevelopment of postural controlluxmee bhattaraiNo ratings yet
- Our Best-Selling Platform Lift: Aritco 7000Document2 pagesOur Best-Selling Platform Lift: Aritco 7000Lê Văn HoạtNo ratings yet
- Physical Pharmaceutics - 1Document15 pagesPhysical Pharmaceutics - 1Odin GODNo ratings yet
- Cellular AberrationDocument8 pagesCellular AberrationRaymund IdicaNo ratings yet
- Altop Industries Limited: Sri Lakshmi Plastic IndustriesDocument3 pagesAltop Industries Limited: Sri Lakshmi Plastic IndustriesSMART T PlacementsNo ratings yet
- Ethernet/Ip: Technical ManualDocument68 pagesEthernet/Ip: Technical ManualDiana HerreraNo ratings yet
- Wlna9ce - Parts ManualDocument24 pagesWlna9ce - Parts Manualsclaveau20No ratings yet
- Nanazoxid 0Document5 pagesNanazoxid 0hnn9gfqzqxNo ratings yet
- The Story That Must Not Be Told by Kavery NambisanDocument6 pagesThe Story That Must Not Be Told by Kavery Nambisanfreelancer800100% (1)
- Comparative Evaluation of Antibacterial Efficacy.9Document6 pagesComparative Evaluation of Antibacterial Efficacy.9Shivani DubeyNo ratings yet
- Aoralscan 2 User Manual+Document8 pagesAoralscan 2 User Manual+MAHESH KULKARNINo ratings yet
- Vegan Mayonnaise - Simple Vegan Blog PDFDocument2 pagesVegan Mayonnaise - Simple Vegan Blog PDFjkoiluNo ratings yet
- Networks 512 CA Test 2Document54 pagesNetworks 512 CA Test 2Thina Siphesihle ZuluNo ratings yet
- LSS Catalogue enDocument68 pagesLSS Catalogue enArzu AliyevaNo ratings yet
- CH 18Document9 pagesCH 18Julie Nix FrazierNo ratings yet
- Masters of Sex 1x01 - PilotDocument66 pagesMasters of Sex 1x01 - PilotettaworNo ratings yet
- Disciplines and Ideas in The Social SciencesDocument18 pagesDisciplines and Ideas in The Social SciencesArjane Grace SullanoNo ratings yet
- Ketone Bodies: DR Anjali SaxenaDocument3 pagesKetone Bodies: DR Anjali Saxenahirendra patelNo ratings yet
- Btech 1 Sem Programming For Problem Solving kcs101 2021Document1 pageBtech 1 Sem Programming For Problem Solving kcs101 2021rishabhchauhan2266No ratings yet
- A Place You Want To Stay at On Your Next TripDocument4 pagesA Place You Want To Stay at On Your Next TripĐoàn Thanh Chúc (FGW HN)No ratings yet
- L A Level Physics A MS Jun 07 Old g481Document88 pagesL A Level Physics A MS Jun 07 Old g481dasha962No ratings yet
- ASU-C-1010 (Receiving Inspection - Bolts, Nuts & Washers For Structural Connections) 14122016Document7 pagesASU-C-1010 (Receiving Inspection - Bolts, Nuts & Washers For Structural Connections) 14122016Bhavanishankar ShettyNo ratings yet
- Annual Day AnchoringDocument5 pagesAnnual Day AnchoringPooja ChuriwalNo ratings yet
- SCP TRPG Cheatsheet Collection v3 SmallerDocument4 pagesSCP TRPG Cheatsheet Collection v3 SmallerNUNUゲイNo ratings yet
- Middle Ear Tumors - Dr. Shivendra PandeyDocument60 pagesMiddle Ear Tumors - Dr. Shivendra PandeyNupur GargNo ratings yet