
MUNI Smysly Sluch
MUNI Smysly Sluch
You might also like
- Mráz - Pitevné CvičeniaDocument196 pagesMráz - Pitevné CvičeniacozysleepingareaNo ratings yet
- LadičkyDocument70 pagesLadičkyLea ShantiNo ratings yet
- 21 Vseob Podnet PocitekDocument52 pages21 Vseob Podnet PocitekKarin GergelyováNo ratings yet
- PL 8 Reflex SmyslyDocument18 pagesPL 8 Reflex SmyslyHonza MikešNo ratings yet
- Smyslová SoustavaDocument9 pagesSmyslová Soustavagabriela.vozenilkovaNo ratings yet
- Nervová SoustavaDocument8 pagesNervová Soustavaklara musalkovaNo ratings yet
- Nervová SoustavaDocument12 pagesNervová Soustavagabriela.vozenilkovaNo ratings yet
- Nervová SoustavaDocument9 pagesNervová Soustavajanmanhal136No ratings yet
- NervovásoustavaDocument13 pagesNervovásoustavaDominika LochmanováNo ratings yet
- Fylogeneze Smyslové Soustavy - 1.částDocument42 pagesFylogeneze Smyslové Soustavy - 1.částBestlolVideo :No ratings yet
- Rehabilitace u lézí PNS (2. LF UK)Document39 pagesRehabilitace u lézí PNS (2. LF UK)kristyna.regentovaNo ratings yet
- ZL - Nervová Soustava - Pracovnã ListDocument5 pagesZL - Nervová Soustava - Pracovnã Listtadus05No ratings yet
- MUNI Ultrazvukova DiagnostikaDocument61 pagesMUNI Ultrazvukova DiagnostikacozysleepingareaNo ratings yet
- Uvod Do Nervove SoustavyDocument15 pagesUvod Do Nervove SoustavyRadim ObchodničekNo ratings yet
- Smyslová Soustava 2Document7 pagesSmyslová Soustava 2MaxNo ratings yet
- Obecka Komplet Výcuc Víc Stejně NenapíšešDocument29 pagesObecka Komplet Výcuc Víc Stejně Nenapíšešepicw059No ratings yet
- 24 - Lidské Smysly A Látkové Řízení Organismu ČlověkaDocument15 pages24 - Lidské Smysly A Látkové Řízení Organismu Člověkahana.hovorkovaNo ratings yet
- Fyziologické Mechanismy PsychikyDocument2 pagesFyziologické Mechanismy PsychikystepanNo ratings yet
- Snímek Obrazovky 2024-03-13 V 12.30.36 Odp.Document9 pagesSnímek Obrazovky 2024-03-13 V 12.30.36 Odp.7f6mkhzchkNo ratings yet
- NERVOVÁ BUŇKA (Neuron) : Periferní DendritDocument24 pagesNERVOVÁ BUŇKA (Neuron) : Periferní DendritDan Anton PetrilaNo ratings yet
- 19 Nervova SoustavaDocument12 pages19 Nervova SoustavaMartin CemerekNo ratings yet
- Úvod Do Fyzikální TerapieDocument147 pagesÚvod Do Fyzikální TerapiemichNo ratings yet
- Fyziologie Svalu A PohybuDocument3 pagesFyziologie Svalu A PohybuBarbora K.No ratings yet
- Neurologie Obecka MisaDocument52 pagesNeurologie Obecka MisaKristINo ratings yet
- Chirurgie 2019 KompletDocument354 pagesChirurgie 2019 KompletAdam SkreckNo ratings yet
- Nervová SoustavaDocument5 pagesNervová SoustavaMagdaléna VíchováNo ratings yet
- 14 15 Smyslove OrganyDocument6 pages14 15 Smyslove OrganyadelkaskramuskaNo ratings yet
- Nervová Soustava 2Document7 pagesNervová Soustava 2beekey95No ratings yet
- RTG Komplet - Uprava JBDocument17 pagesRTG Komplet - Uprava JBKristINo ratings yet
- Nervová SoustavaDocument6 pagesNervová Soustavaema.varejkovaNo ratings yet
- Nervno Tkivo I Nervni SistemDocument65 pagesNervno Tkivo I Nervni Sistemmilana95prlinaNo ratings yet
- Infrazvuk A UltrazvukDocument7 pagesInfrazvuk A UltrazvukVeru CiencialováNo ratings yet
- Základy EegDocument53 pagesZáklady EegJohn ConnorNo ratings yet
- Nervová Soustava A SmyslyDocument14 pagesNervová Soustava A SmyslyLucieNo ratings yet
- Smysly - Čich A ChuťDocument4 pagesSmysly - Čich A ChuťPoli PopováNo ratings yet
- Snímek Obrazovky 2024-02-22 V 1.37.52 Odp.Document14 pagesSnímek Obrazovky 2024-02-22 V 1.37.52 Odp.7f6mkhzchkNo ratings yet
- FT PrezentaceDocument13 pagesFT Prezentacejahn vahnNo ratings yet
- Nervova Soustava Zaostreno Na Neuron 1631947055Document3 pagesNervova Soustava Zaostreno Na Neuron 1631947055TerezaNo ratings yet
- Elektromagnetické Záření (1-4)Document9 pagesElektromagnetické Záření (1-4)lucilleNo ratings yet
- Psychologie, Věda o Chování A Prožívání ČlověkaDocument3 pagesPsychologie, Věda o Chování A Prožívání ČlověkamajaNo ratings yet
- O ZVUKU Pro StudentyDocument14 pagesO ZVUKU Pro StudentyKateřina LandováNo ratings yet
- MUNI-rtg-CTDocument53 pagesMUNI-rtg-CTcozysleepingareaNo ratings yet
- Obecná NeurologieDocument86 pagesObecná Neurologiepw6g8pdbtdNo ratings yet
- 02 BPC-UIN Inteligence 2023Document52 pages02 BPC-UIN Inteligence 2023jan prokopNo ratings yet
- 00 FT postupy v klinických oborechDocument20 pages00 FT postupy v klinických oborechkristyna.regentovaNo ratings yet
- CZ - SIS Study - Pain ManagementDocument6 pagesCZ - SIS Study - Pain Managementf.mariajdbNo ratings yet
- AkustikaDocument23 pagesAkustikamalekNo ratings yet
- Vypracované Otázky K ORL TestuDocument31 pagesVypracované Otázky K ORL TestutheshadowdwellerNo ratings yet
- Smyslová SoustavaDocument43 pagesSmyslová SoustavakaterinasumikovaNo ratings yet
- Nervnotkivosajt PDFDocument76 pagesNervnotkivosajt PDFmolla e embelNo ratings yet
- Article 4Document4 pagesArticle 4janaospalaNo ratings yet
- Obecná PsychologieDocument7 pagesObecná PsychologieVeru ZelinkováNo ratings yet
- Histologické Techniky: A) Světelná Mikroskopie B) Fluorescenční Mikroskopie C) Elektronová Mikroskopie D) AutoradiografieDocument32 pagesHistologické Techniky: A) Světelná Mikroskopie B) Fluorescenční Mikroskopie C) Elektronová Mikroskopie D) AutoradiografieDan Anton PetrilaNo ratings yet
- RovnovahaDocument2 pagesRovnovahaPoli PopováNo ratings yet
- PrázdnáDocument6 pagesPrázdnáVeronikaNo ratings yet
- Histologie 1 - Úvod, EpitelyDocument2 pagesHistologie 1 - Úvod, EpitelyJiří KoutNo ratings yet
- Bezkontaktná MasážDocument129 pagesBezkontaktná MasážJano DančejNo ratings yet
- Nervová SoustavaDocument2 pagesNervová SoustavaLucieNo ratings yet
- Jak Poznat, Že EKG Je Abnormální A Co Nám Zobrazuje Standardní EKG ZáznamDocument5 pagesJak Poznat, Že EKG Je Abnormální A Co Nám Zobrazuje Standardní EKG ZáznambalounovaNo ratings yet
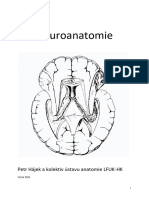
- Neuroanatomie: Petr Hájek A Kolektiv Ústavu Anatomie LFUK-HKDocument245 pagesNeuroanatomie: Petr Hájek A Kolektiv Ústavu Anatomie LFUK-HKLucie VokálováNo ratings yet
- MUNI-oko (1)Document45 pagesMUNI-oko (1)cozysleepingareaNo ratings yet
- MUNI-nuklearni Medicina a Radioterapie-21Document59 pagesMUNI-nuklearni Medicina a Radioterapie-21cozysleepingareaNo ratings yet
- MUNI Ultrazvukova DiagnostikaDocument61 pagesMUNI Ultrazvukova DiagnostikacozysleepingareaNo ratings yet
- MUNI Termodynamika Principy 21Document17 pagesMUNI Termodynamika Principy 21cozysleepingareaNo ratings yet
- MUNI-rtg-CTDocument53 pagesMUNI-rtg-CTcozysleepingareaNo ratings yet
- MUNI MRI UpravaMDspoznamkami 21Document34 pagesMUNI MRI UpravaMDspoznamkami 21cozysleepingareaNo ratings yet
































































Download as pptx, pdf, or txt
You might also like
- Mráz - Pitevné CvičeniaDocument196 pagesMráz - Pitevné CvičeniacozysleepingareaNo ratings yet
- LadičkyDocument70 pagesLadičkyLea ShantiNo ratings yet
- 21 Vseob Podnet PocitekDocument52 pages21 Vseob Podnet PocitekKarin GergelyováNo ratings yet
- PL 8 Reflex SmyslyDocument18 pagesPL 8 Reflex SmyslyHonza MikešNo ratings yet
- Smyslová SoustavaDocument9 pagesSmyslová Soustavagabriela.vozenilkovaNo ratings yet
- Nervová SoustavaDocument8 pagesNervová Soustavaklara musalkovaNo ratings yet
- Nervová SoustavaDocument12 pagesNervová Soustavagabriela.vozenilkovaNo ratings yet
- Nervová SoustavaDocument9 pagesNervová Soustavajanmanhal136No ratings yet
- NervovásoustavaDocument13 pagesNervovásoustavaDominika LochmanováNo ratings yet
- Fylogeneze Smyslové Soustavy - 1.částDocument42 pagesFylogeneze Smyslové Soustavy - 1.částBestlolVideo :No ratings yet
- Rehabilitace u lézí PNS (2. LF UK)Document39 pagesRehabilitace u lézí PNS (2. LF UK)kristyna.regentovaNo ratings yet
- ZL - Nervová Soustava - Pracovnã ListDocument5 pagesZL - Nervová Soustava - Pracovnã Listtadus05No ratings yet
- MUNI Ultrazvukova DiagnostikaDocument61 pagesMUNI Ultrazvukova DiagnostikacozysleepingareaNo ratings yet
- Uvod Do Nervove SoustavyDocument15 pagesUvod Do Nervove SoustavyRadim ObchodničekNo ratings yet
- Smyslová Soustava 2Document7 pagesSmyslová Soustava 2MaxNo ratings yet
- Obecka Komplet Výcuc Víc Stejně NenapíšešDocument29 pagesObecka Komplet Výcuc Víc Stejně Nenapíšešepicw059No ratings yet
- 24 - Lidské Smysly A Látkové Řízení Organismu ČlověkaDocument15 pages24 - Lidské Smysly A Látkové Řízení Organismu Člověkahana.hovorkovaNo ratings yet
- Fyziologické Mechanismy PsychikyDocument2 pagesFyziologické Mechanismy PsychikystepanNo ratings yet
- Snímek Obrazovky 2024-03-13 V 12.30.36 Odp.Document9 pagesSnímek Obrazovky 2024-03-13 V 12.30.36 Odp.7f6mkhzchkNo ratings yet
- NERVOVÁ BUŇKA (Neuron) : Periferní DendritDocument24 pagesNERVOVÁ BUŇKA (Neuron) : Periferní DendritDan Anton PetrilaNo ratings yet
- 19 Nervova SoustavaDocument12 pages19 Nervova SoustavaMartin CemerekNo ratings yet
- Úvod Do Fyzikální TerapieDocument147 pagesÚvod Do Fyzikální TerapiemichNo ratings yet
- Fyziologie Svalu A PohybuDocument3 pagesFyziologie Svalu A PohybuBarbora K.No ratings yet
- Neurologie Obecka MisaDocument52 pagesNeurologie Obecka MisaKristINo ratings yet
- Chirurgie 2019 KompletDocument354 pagesChirurgie 2019 KompletAdam SkreckNo ratings yet
- Nervová SoustavaDocument5 pagesNervová SoustavaMagdaléna VíchováNo ratings yet
- 14 15 Smyslove OrganyDocument6 pages14 15 Smyslove OrganyadelkaskramuskaNo ratings yet
- Nervová Soustava 2Document7 pagesNervová Soustava 2beekey95No ratings yet
- RTG Komplet - Uprava JBDocument17 pagesRTG Komplet - Uprava JBKristINo ratings yet
- Nervová SoustavaDocument6 pagesNervová Soustavaema.varejkovaNo ratings yet
- Nervno Tkivo I Nervni SistemDocument65 pagesNervno Tkivo I Nervni Sistemmilana95prlinaNo ratings yet
- Infrazvuk A UltrazvukDocument7 pagesInfrazvuk A UltrazvukVeru CiencialováNo ratings yet
- Základy EegDocument53 pagesZáklady EegJohn ConnorNo ratings yet
- Nervová Soustava A SmyslyDocument14 pagesNervová Soustava A SmyslyLucieNo ratings yet
- Smysly - Čich A ChuťDocument4 pagesSmysly - Čich A ChuťPoli PopováNo ratings yet
- Snímek Obrazovky 2024-02-22 V 1.37.52 Odp.Document14 pagesSnímek Obrazovky 2024-02-22 V 1.37.52 Odp.7f6mkhzchkNo ratings yet
- FT PrezentaceDocument13 pagesFT Prezentacejahn vahnNo ratings yet
- Nervova Soustava Zaostreno Na Neuron 1631947055Document3 pagesNervova Soustava Zaostreno Na Neuron 1631947055TerezaNo ratings yet
- Elektromagnetické Záření (1-4)Document9 pagesElektromagnetické Záření (1-4)lucilleNo ratings yet
- Psychologie, Věda o Chování A Prožívání ČlověkaDocument3 pagesPsychologie, Věda o Chování A Prožívání ČlověkamajaNo ratings yet
- O ZVUKU Pro StudentyDocument14 pagesO ZVUKU Pro StudentyKateřina LandováNo ratings yet
- MUNI-rtg-CTDocument53 pagesMUNI-rtg-CTcozysleepingareaNo ratings yet
- Obecná NeurologieDocument86 pagesObecná Neurologiepw6g8pdbtdNo ratings yet
- 02 BPC-UIN Inteligence 2023Document52 pages02 BPC-UIN Inteligence 2023jan prokopNo ratings yet
- 00 FT postupy v klinických oborechDocument20 pages00 FT postupy v klinických oborechkristyna.regentovaNo ratings yet
- CZ - SIS Study - Pain ManagementDocument6 pagesCZ - SIS Study - Pain Managementf.mariajdbNo ratings yet
- AkustikaDocument23 pagesAkustikamalekNo ratings yet
- Vypracované Otázky K ORL TestuDocument31 pagesVypracované Otázky K ORL TestutheshadowdwellerNo ratings yet
- Smyslová SoustavaDocument43 pagesSmyslová SoustavakaterinasumikovaNo ratings yet
- Nervnotkivosajt PDFDocument76 pagesNervnotkivosajt PDFmolla e embelNo ratings yet
- Article 4Document4 pagesArticle 4janaospalaNo ratings yet
- Obecná PsychologieDocument7 pagesObecná PsychologieVeru ZelinkováNo ratings yet
- Histologické Techniky: A) Světelná Mikroskopie B) Fluorescenční Mikroskopie C) Elektronová Mikroskopie D) AutoradiografieDocument32 pagesHistologické Techniky: A) Světelná Mikroskopie B) Fluorescenční Mikroskopie C) Elektronová Mikroskopie D) AutoradiografieDan Anton PetrilaNo ratings yet
- RovnovahaDocument2 pagesRovnovahaPoli PopováNo ratings yet
- PrázdnáDocument6 pagesPrázdnáVeronikaNo ratings yet
- Histologie 1 - Úvod, EpitelyDocument2 pagesHistologie 1 - Úvod, EpitelyJiří KoutNo ratings yet
- Bezkontaktná MasážDocument129 pagesBezkontaktná MasážJano DančejNo ratings yet
- Nervová SoustavaDocument2 pagesNervová SoustavaLucieNo ratings yet
- Jak Poznat, Že EKG Je Abnormální A Co Nám Zobrazuje Standardní EKG ZáznamDocument5 pagesJak Poznat, Že EKG Je Abnormální A Co Nám Zobrazuje Standardní EKG ZáznambalounovaNo ratings yet
- Neuroanatomie: Petr Hájek A Kolektiv Ústavu Anatomie LFUK-HKDocument245 pagesNeuroanatomie: Petr Hájek A Kolektiv Ústavu Anatomie LFUK-HKLucie VokálováNo ratings yet
- MUNI-oko (1)Document45 pagesMUNI-oko (1)cozysleepingareaNo ratings yet
- MUNI-nuklearni Medicina a Radioterapie-21Document59 pagesMUNI-nuklearni Medicina a Radioterapie-21cozysleepingareaNo ratings yet
- MUNI Ultrazvukova DiagnostikaDocument61 pagesMUNI Ultrazvukova DiagnostikacozysleepingareaNo ratings yet
- MUNI Termodynamika Principy 21Document17 pagesMUNI Termodynamika Principy 21cozysleepingareaNo ratings yet
- MUNI-rtg-CTDocument53 pagesMUNI-rtg-CTcozysleepingareaNo ratings yet
- MUNI MRI UpravaMDspoznamkami 21Document34 pagesMUNI MRI UpravaMDspoznamkami 21cozysleepingareaNo ratings yet